

Human RNase H1 is associated with protein P32 and is involved in mitochondrial pre-rRNA processing
- PMID: 23990920
- PMCID: PMC3750045
- DOI: 10.1371/journal.pone.0071006
Human RNase H1 is associated with protein P32 and is involved in mitochondrial pre-rRNA processing
Abstract
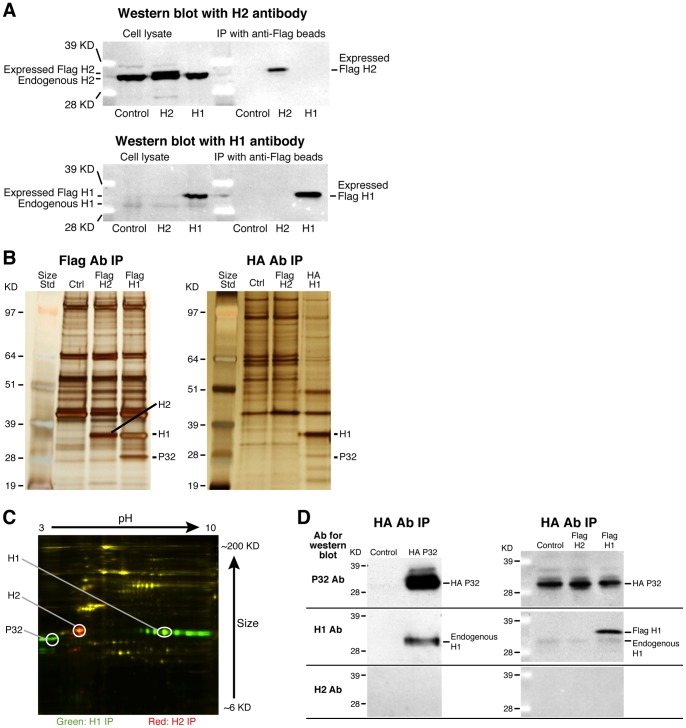
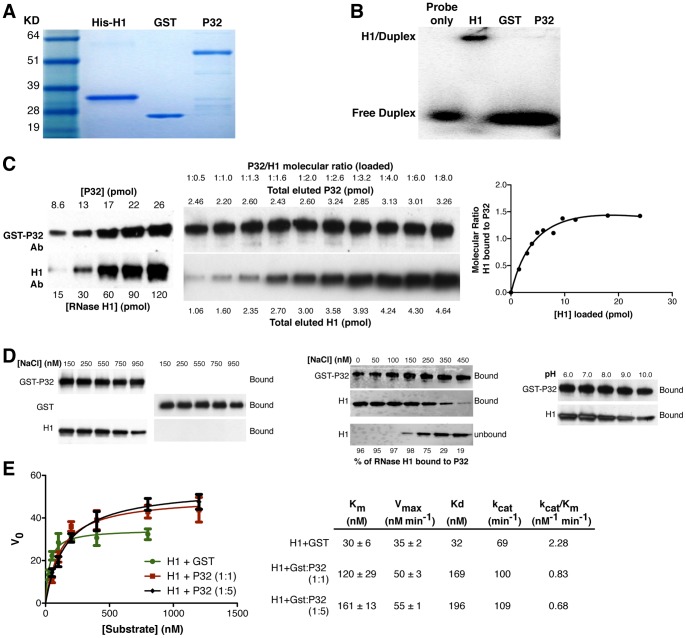
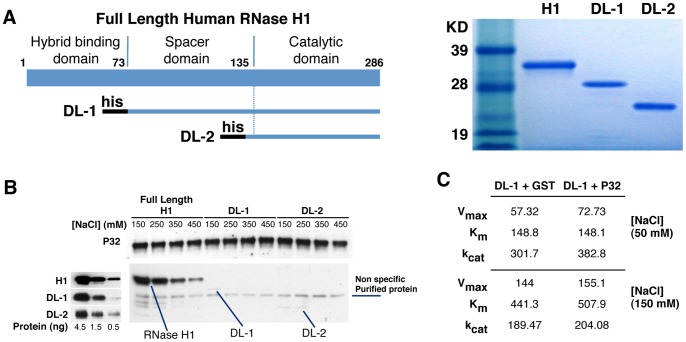
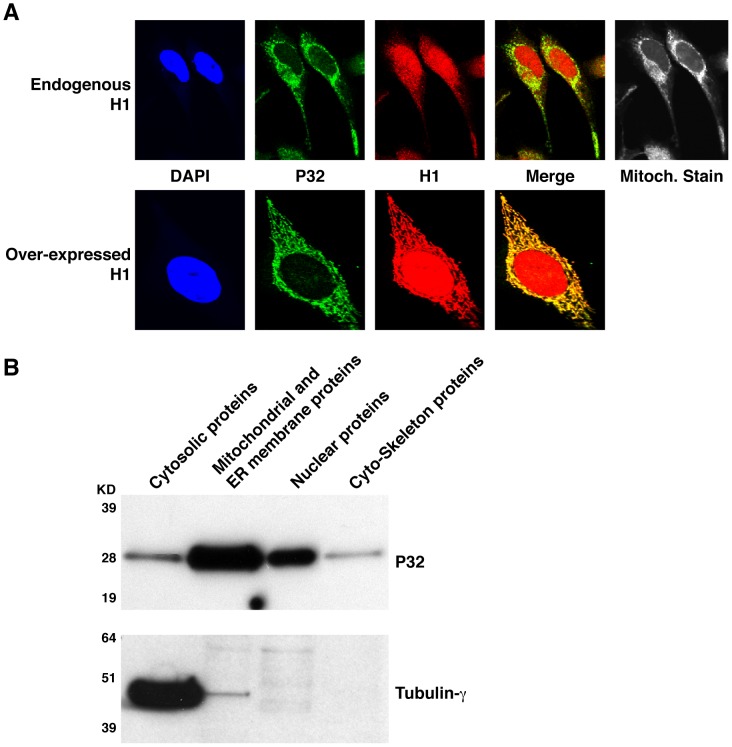
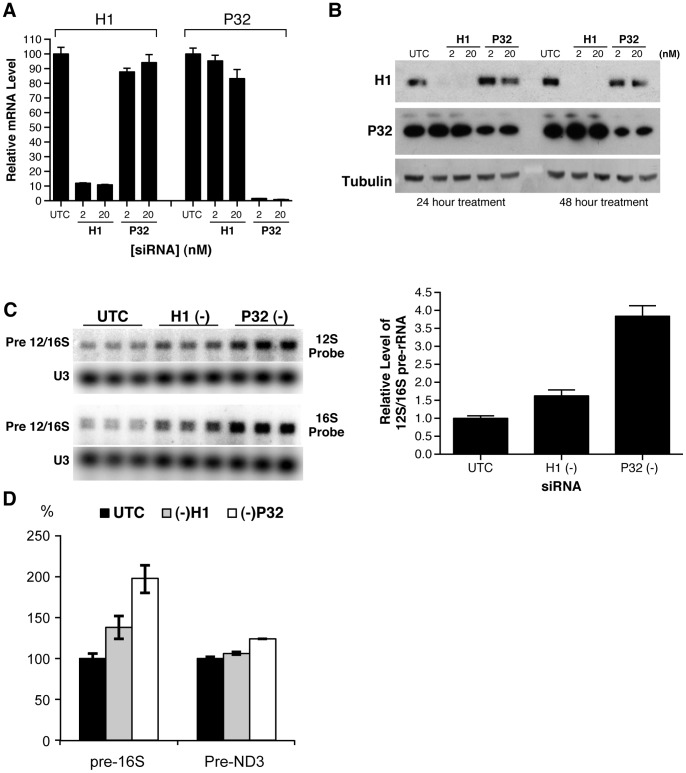
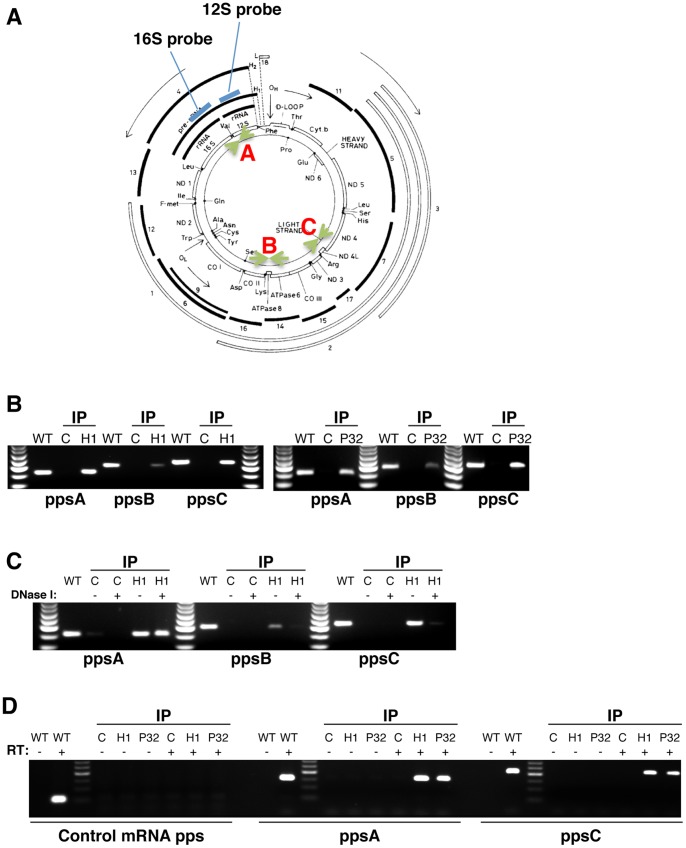
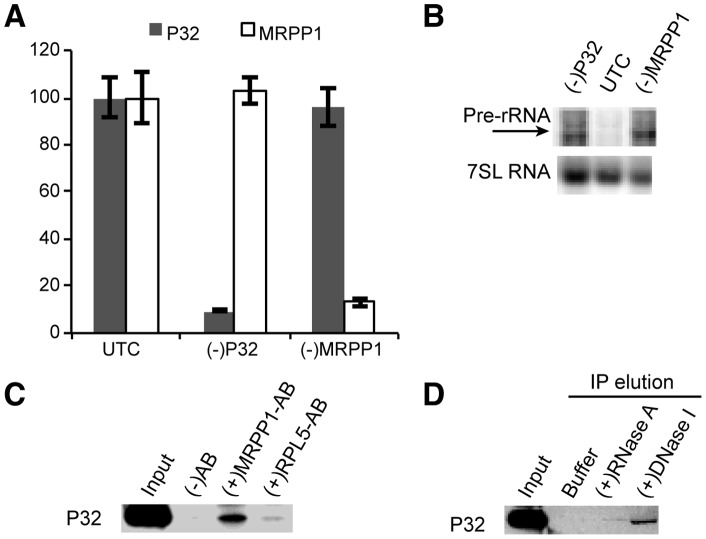
Mammalian RNase H1 has been implicated in mitochondrial DNA replication and RNA processing and is required for embryonic development. We identified the mitochondrial protein P32 that binds specifically to human RNase H1, but not human RNase H2. P32 binds human RNase H1 via the hybrid-binding domain of the enzyme at an approximately 1∶1 ratio. P32 enhanced the cleavage activity of RNase H1 by reducing the affinity of the enzyme for the heteroduplex substrate and enhancing turnover, but had no effect on the cleavage pattern. RNase H1 and P32 were partially co-localized in mitochondria and reduction of P32 or RNase H1 levels resulted in accumulation of mitochondrial pre ribosomal RNA [12S/16S] in HeLa cells. P32 also co-immunoprecipitated with MRPP1, a mitochondrial RNase P protein required for mitochondrial pre-rRNA processing. The P32-RNase H1 complex was shown to physically interact with mitochondrial DNA and pre-rRNA. These results expand the potential roles for RNase H1 to include assuring proper transcription and processing of guanosine-cytosine rich pre-ribosomal RNA in mitochondria. Further, the results identify P32 as a member of the 'RNase H1 degradosome' and the key P32 enhances the enzymatic efficiency of human RNase H1.
Conflict of interest statement
Figures
References
-
- Stein H, Hausen P (1969) Enzyme from calf thymus degrading the RNA moiety of DNA-RNA Hybrids: effect on DNA-dependent RNA polymerase. Science, 166, 393–395. - PubMed
-
- Büsen (1980) Purification, subunit structure, and serologicai analysis of calf thymus ribonuclease H I. The Journal of biological chemistry, 255, 9434–9443. - PubMed
-
- Rong YW, Carl PL (1990) On the molecular weight and subunit composition of calf thymus ribonuclease H1. Biochemistry, 29, 383–389. - PubMed
-
- Itaya M, McKelvin D, Chatterjie SK, Crouch RJ (1991) Selective cloning of genes encoding RNase H from Salmonella typhimurium, Saccharomyces cerevisiae and Escherichia coli rnh mutant. Molecular & general genetics: MGG, 227, 438–445. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
