

Porphyromonas gingivalis promotes invasion of oral squamous cell carcinoma through induction of proMMP9 and its activation
- PMID: 23991831
- PMCID: PMC3939075
- DOI: 10.1111/cmi.12211
Porphyromonas gingivalis promotes invasion of oral squamous cell carcinoma through induction of proMMP9 and its activation
Abstract
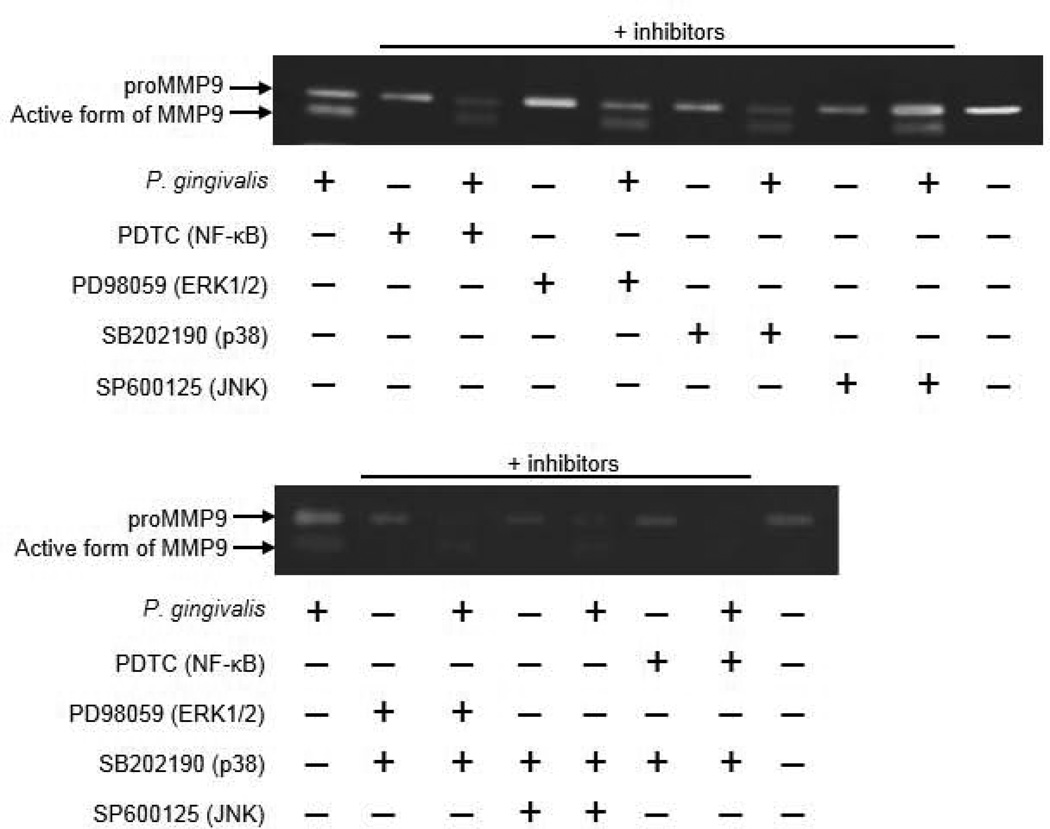
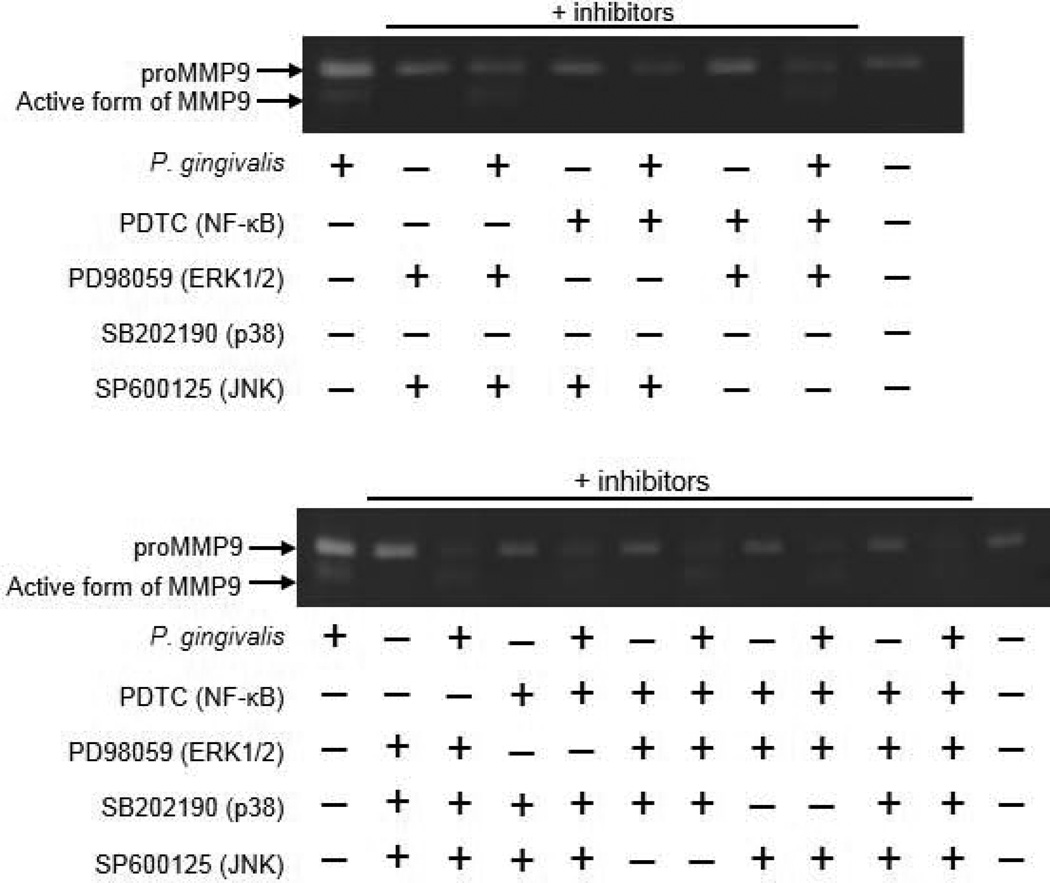
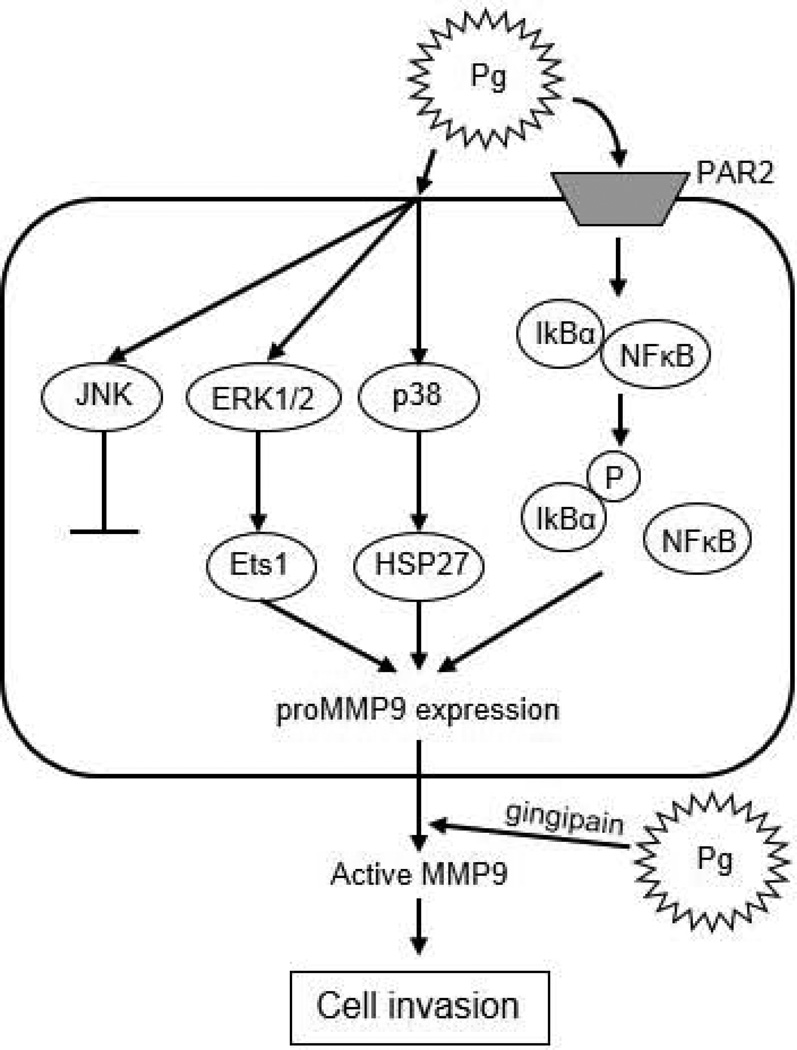
Recent epidemiological studies have revealed a significant association between periodontitis and oral squamous cell carcinoma (OSCC). Furthermore, matrix metalloproteinase 9 (MMP9) is implicated in the invasion and metastasis of tumour cells. We examined the involvement of Porphyromonas gingivalis, a periodontal pathogen, in OSCC invasion through induced expression of proMMP and its activation. proMMP9 was continuously secreted from carcinoma SAS cells, while P. gingivalis infection increased proenzyme expression and subsequently processed it to active MMP9 in culture supernatant, which enhanced cellular invasion. In contrast, Fusobacterium nucleatum, another periodontal organism, failed to demonstrate such activities. The effects of P. gingivalis were observed with highly invasive cells, but not with the low invasivetype. P. gingivalis also stimulated proteinase-activated receptor 2 (PAR2) and enhanced proMMP9 expression, which promoted cellular invasion. P. gingivalis mutants deficient in gingipain proteases failed to activate MMP9. Infected SAS cells exhibited activation of ERK1/2, p38, and NF-kB, and their inhibitors diminished both proMMP9-overexpression and cellular invasion. Together, our results show that P. gingivalis activates the ERK1/2-Ets1, p38/HSP27, and PAR2/NF-kB pathways to induce proMMP9 expression, after which the proenzyme is activated by gingipains to promote cellular invasion of OSCC cell lines. These findings suggest a novel mechanism of progression and metastasis of OSCC associated with periodontitis.
© 2013 John Wiley & Sons Ltd.
Figures










References
-
- Andrian E, Mostefaoui Y, Rouabhia M, Grenier D. Regulation of matrix metalloproteinases and tissue inhibitors of matrix metalloproteinases by Porphyromonas gingivalis in an engineered human oral mucosa model. J Cell Physiol. 2007;211:56–62. - PubMed
-
- Baker EA, Leaper DJ, Hayter JP, Dickenson AJ. Plasminogen activator system in oral squamous cell carcinoma. Br J Oral Maxillofac Surg. 2007;45:623–627. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
