

Identification of long-lived proteins reveals exceptional stability of essential cellular structures
- PMID: 23993091
- PMCID: PMC3788602
- DOI: 10.1016/j.cell.2013.07.037
Identification of long-lived proteins reveals exceptional stability of essential cellular structures
Abstract
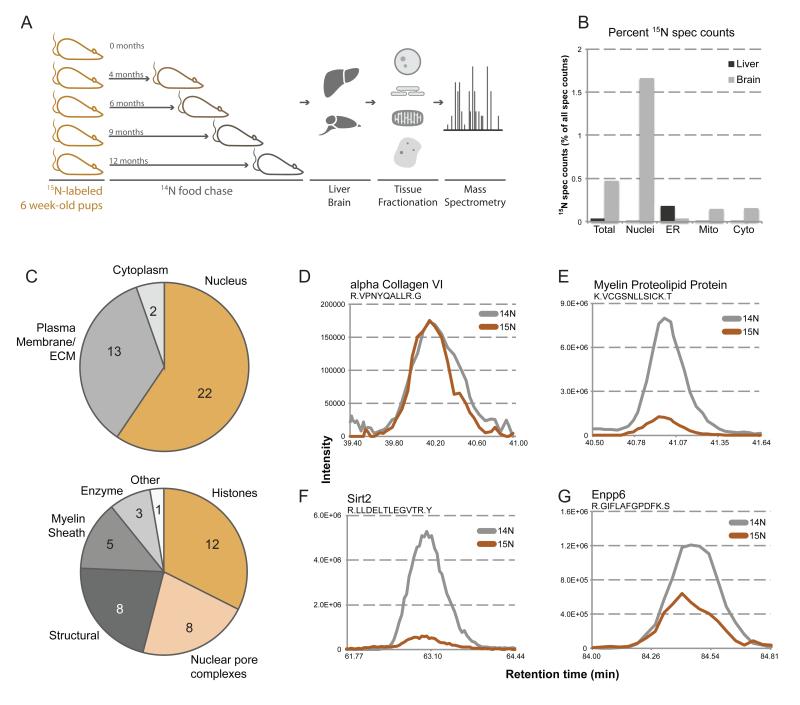
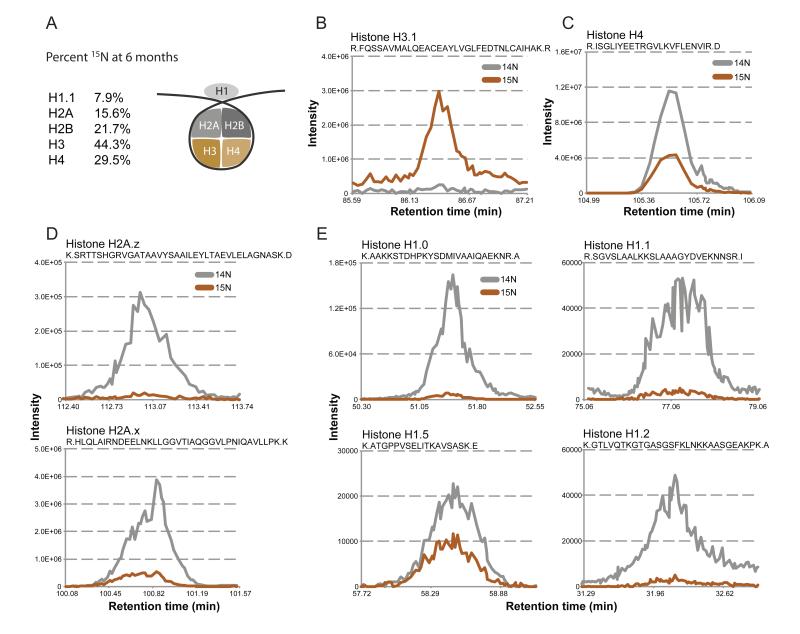
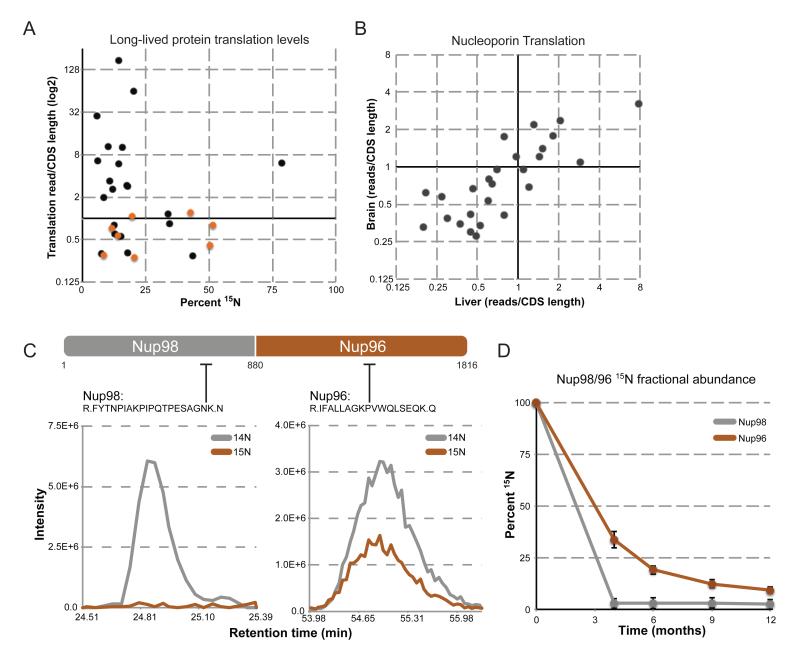
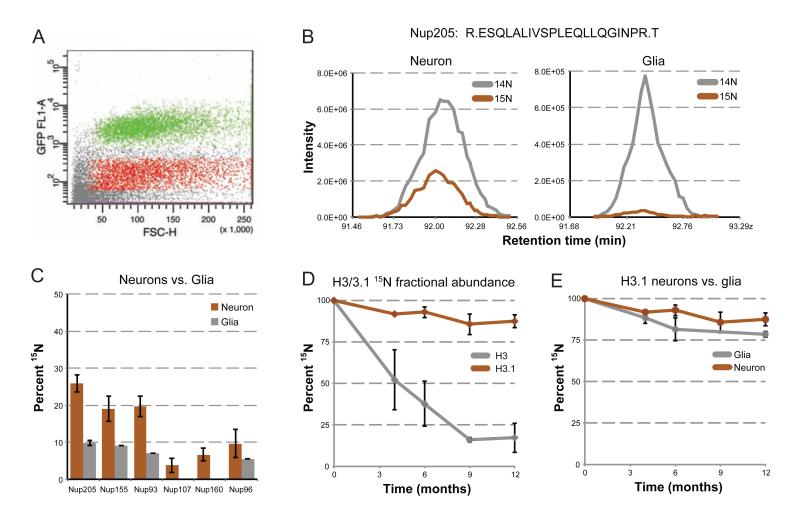
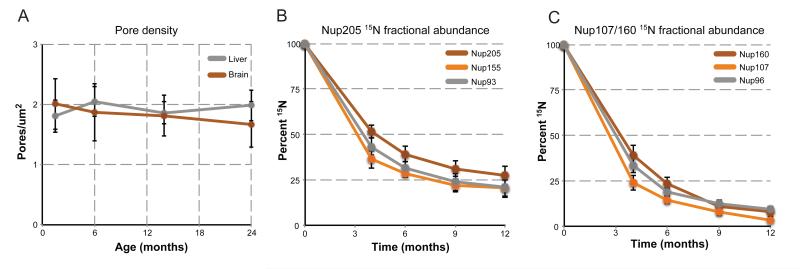
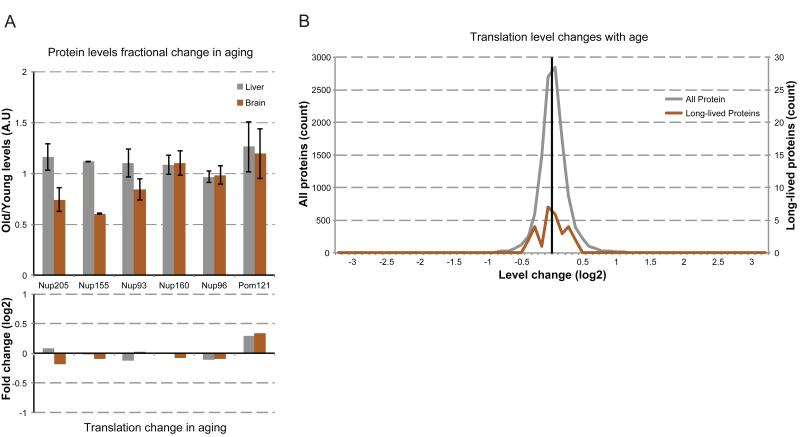
Intracellular proteins with long lifespans have recently been linked to age-dependent defects, ranging from decreased fertility to the functional decline of neurons. Why long-lived proteins exist in metabolically active cellular environments and how they are maintained over time remains poorly understood. Here, we provide a system-wide identification of proteins with exceptional lifespans in the rat brain. These proteins are inefficiently replenished despite being translated robustly throughout adulthood. Using nucleoporins as a paradigm for long-term protein persistence, we found that nuclear pore complexes (NPCs) are maintained over a cell's life through slow but finite exchange of even its most stable subcomplexes. This maintenance is limited, however, as some nucleoporin levels decrease during aging, providing a rationale for the previously observed age-dependent deterioration of NPC function. Our identification of a long-lived proteome reveals cellular components that are at increased risk for damage accumulation, linking long-term protein persistence to the cellular aging process. PAPERCLIP:
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures
References
-
- Ahmad K, Henikoff S. The histone variant H3.3 marks active chromatin by replication-independent nucleosome assembly. Molecular cell. 2002;9:1191–1200. - PubMed
-
- Arber N, Zajicek G, Ariel I. The streaming liver. II. Hepatocyte life history. Liver. 1988;8:80–87. - PubMed
-
- Bassnett S. Lens organelle degradation. Experimental eye research. 2002;74:1–6. - PubMed
