

Probing the selectivity and protein·protein interactions of a nonreducing fungal polyketide synthase using mechanism-based crosslinkers
- PMID: 23993461
- PMCID: PMC3796367
- DOI: 10.1016/j.chembiol.2013.07.012
Probing the selectivity and protein·protein interactions of a nonreducing fungal polyketide synthase using mechanism-based crosslinkers
Erratum in
- Chem Biol. 2014 Jul 17;21(7):913. Haushalter, Bob [corrected to Haushalter, Robert W]; Vagstad, Anna [corrected to Vagstad, Anna L]
Abstract
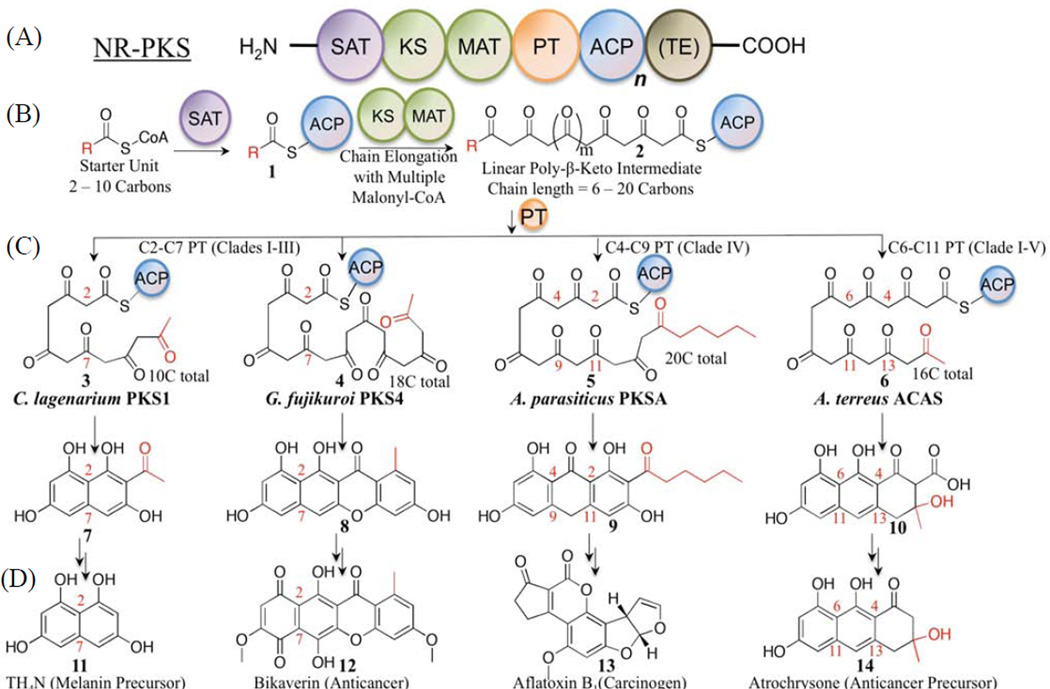
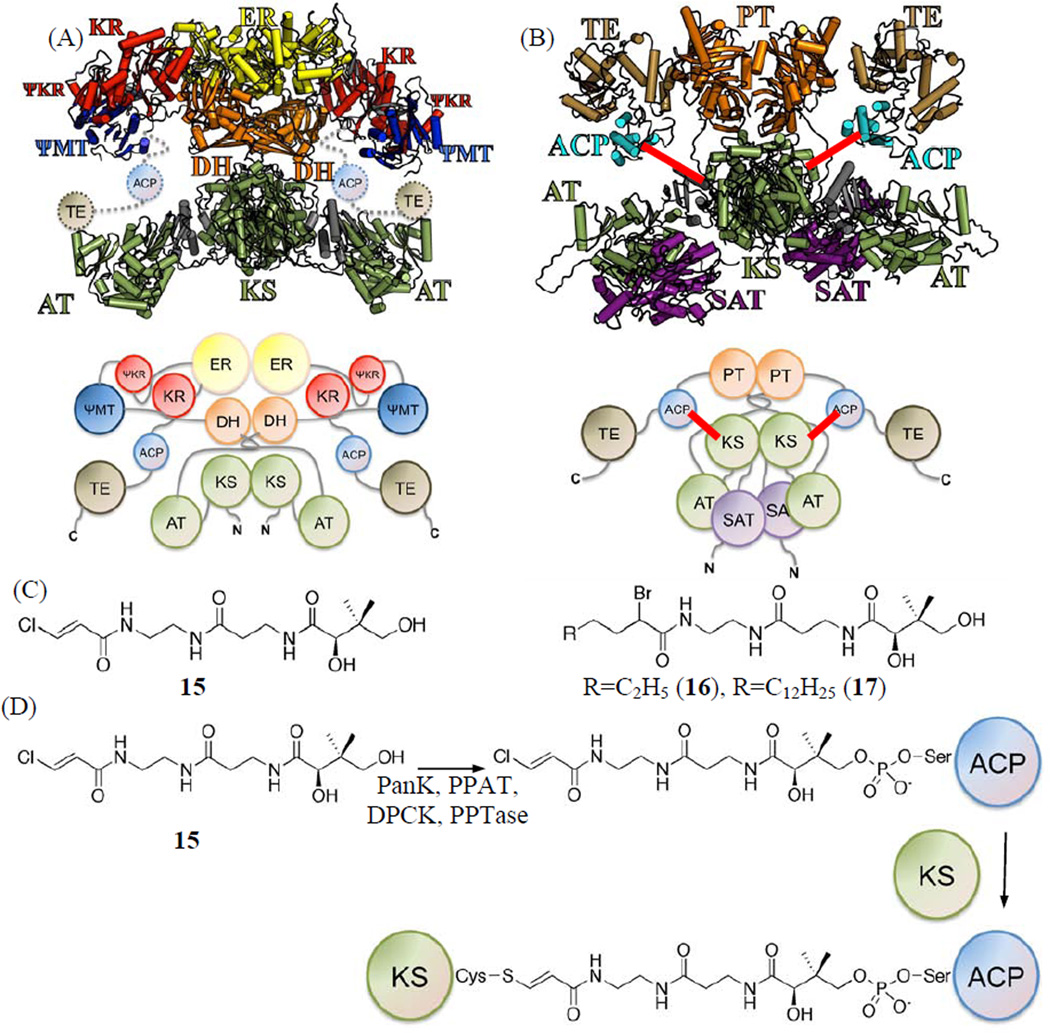
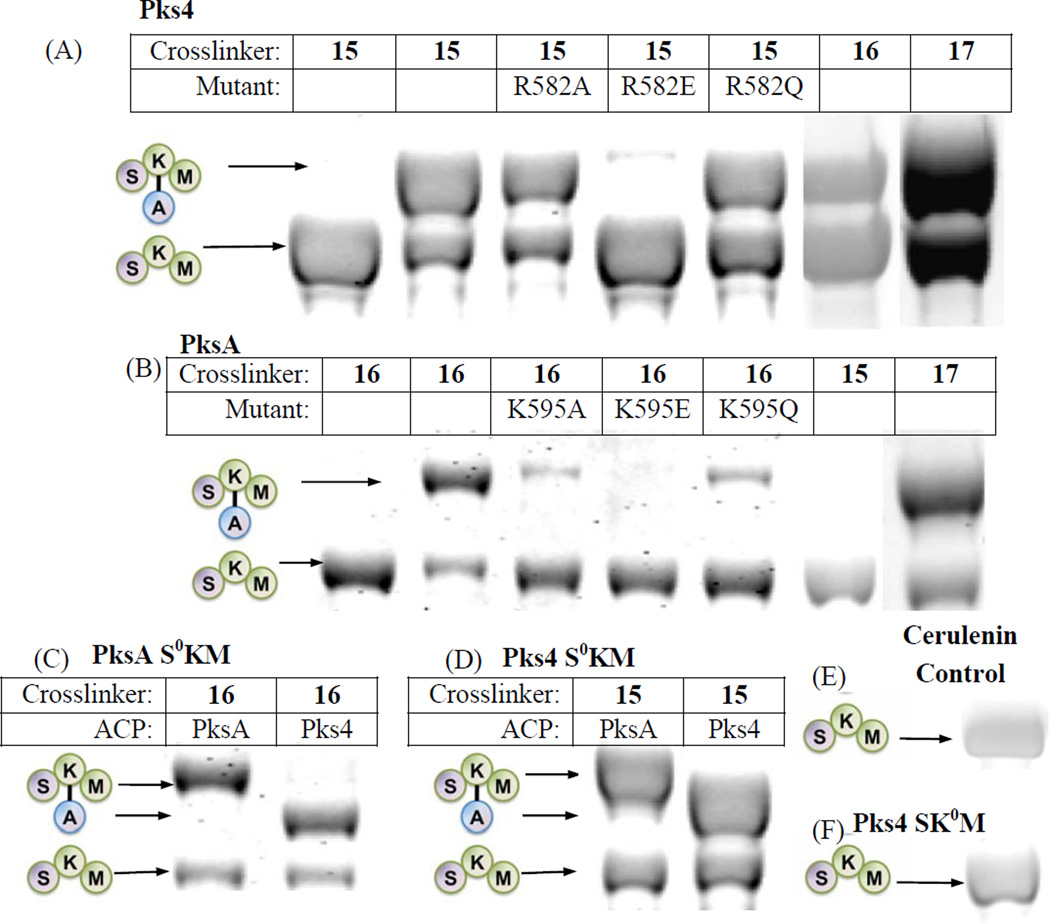
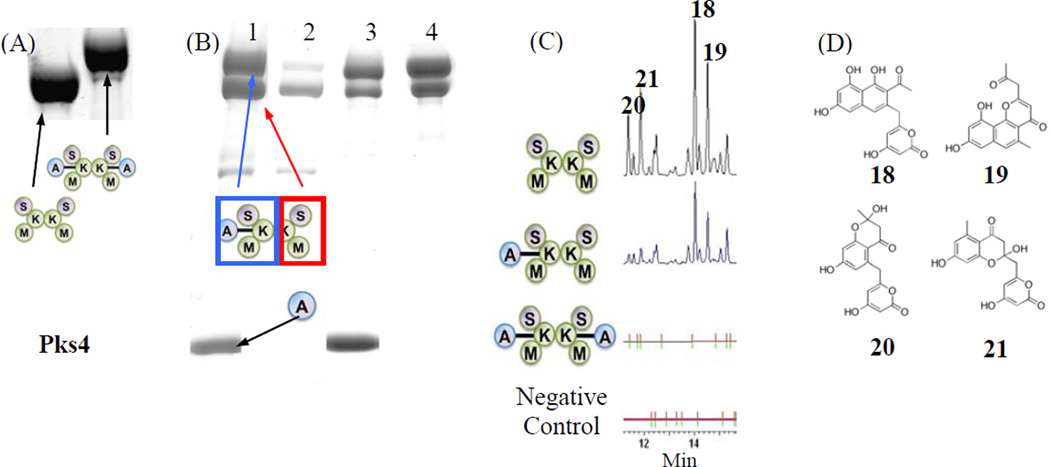
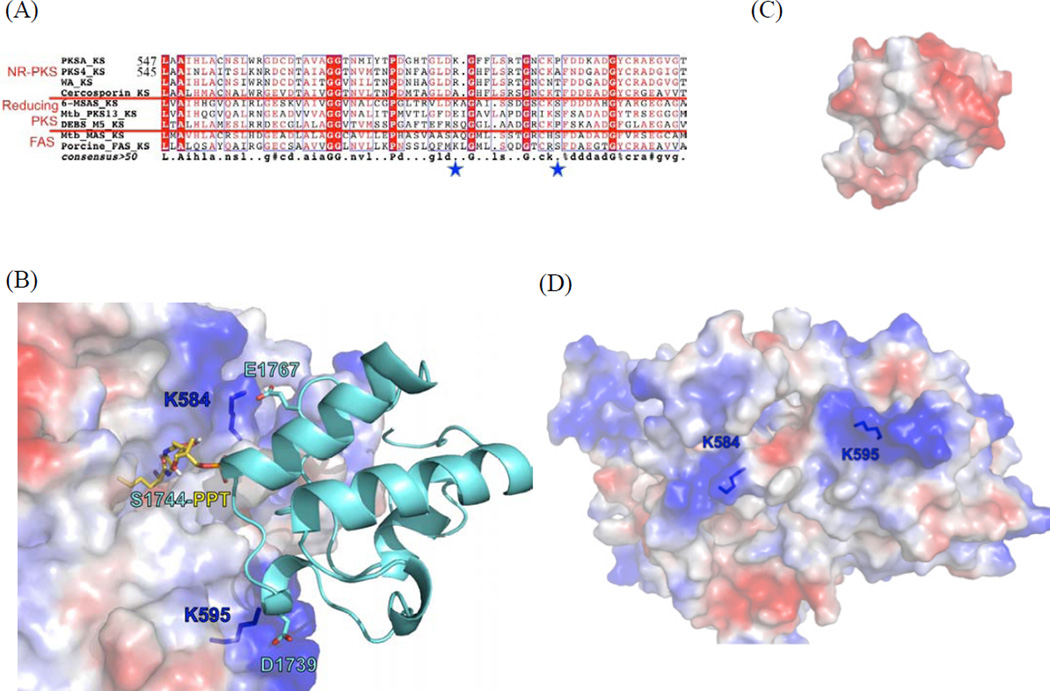
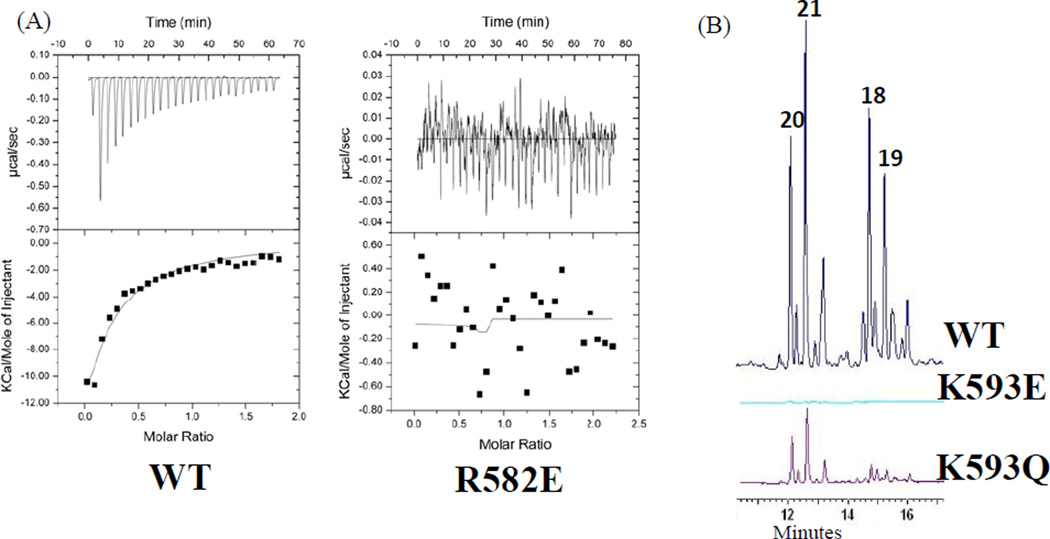
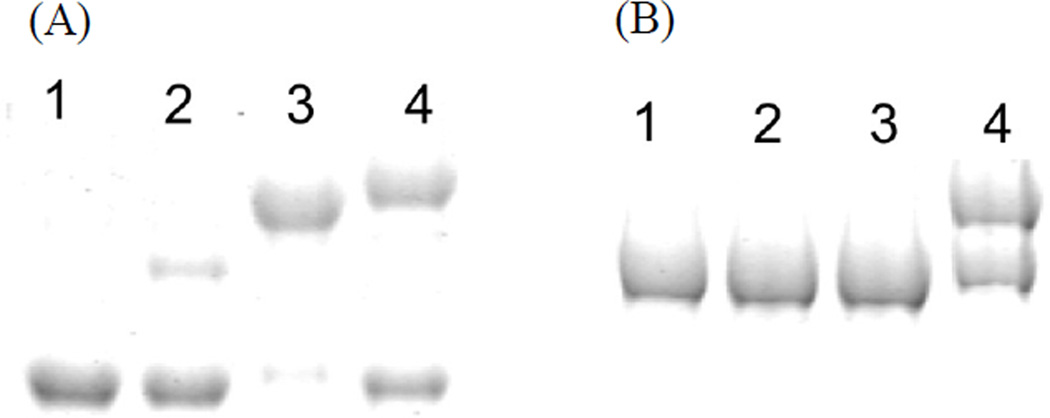
Protein·protein interactions, which often involve interactions among an acyl carrier protein (ACP) and ACP partner enzymes, are important for coordinating polyketide biosynthesis. However, the nature of such interactions is not well understood, especially in the fungal nonreducing polyketide synthases (NR-PKSs) that biosynthesize toxic and pharmaceutically important polyketides. Here, we employ mechanism-based crosslinkers to successfully probe ACP and ketosynthase (KS) domain interactions in NR-PKSs. We found that crosslinking efficiency is closely correlated with the strength of ACP·KS interactions and that KS demonstrates strong starter unit selectivity. We further identified positively charged surface residues by KS mutagenesis, which mediates key interactions with the negatively charged ACP surface. Such complementary/matching contact pairs can serve as "adapter surfaces" for future efforts to generate new polyketides using NR-PKSs.
Copyright © 2013 Elsevier Ltd. All rights reserved.
Figures
Comment in
-
Probing interactions in fungal PKS.Chem Biol. 2013 Sep 19;20(9):1089-91. doi: 10.1016/j.chembiol.2013.09.001. Chem Biol. 2013. PMID: 24054181
References
-
- Cox RJ. Polyketides, proteins and genes in fungi: programmed nano-machines begin to reveal their secrets. Org Biomol Chem. 2007;5:2010–2026. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
