

Extrathymic Aire-expressing cells are a distinct bone marrow-derived population that induce functional inactivation of CD4⁺ T cells
- PMID: 23993652
- PMCID: PMC3804105
- DOI: 10.1016/j.immuni.2013.08.005
Extrathymic Aire-expressing cells are a distinct bone marrow-derived population that induce functional inactivation of CD4⁺ T cells
Abstract
The autoimmune regulator (Aire) is essential for prevention of autoimmunity; its role is best understood in the thymus, where it promotes self-tolerance through tissue-specific antigen (TSA) expression. Recently, extrathymic Aire-expressing cells (eTACs) have been described in murine secondary lymphoid organs, but the identity of such cells and their role in immune tolerance remains unclear. Here we have shown that eTACs are a discrete major histocompatibility complex class II (MHC II)(hi), CD80(lo), CD86(lo), epithelial cell adhesion molecule (EpCAM)(hi), CD45(lo) bone marrow-derived peripheral antigen-presenting cell (APC) population. We also have demonstrated that eTACs can functionally inactivate CD4⁺ T cells through a mechanism that does not require regulatory T cells (Treg) and is resistant to innate inflammatory stimuli. Together, these findings further define eTACs as a distinct tolerogenic cell population in secondary lymphoid organs.
Copyright © 2013 Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
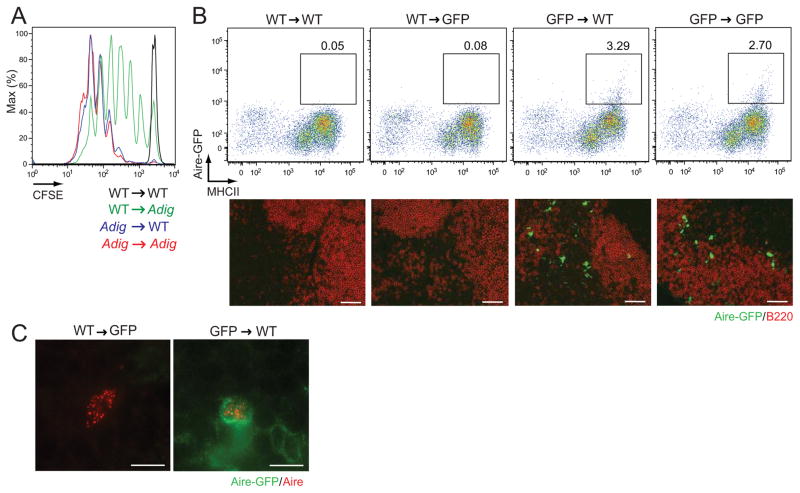
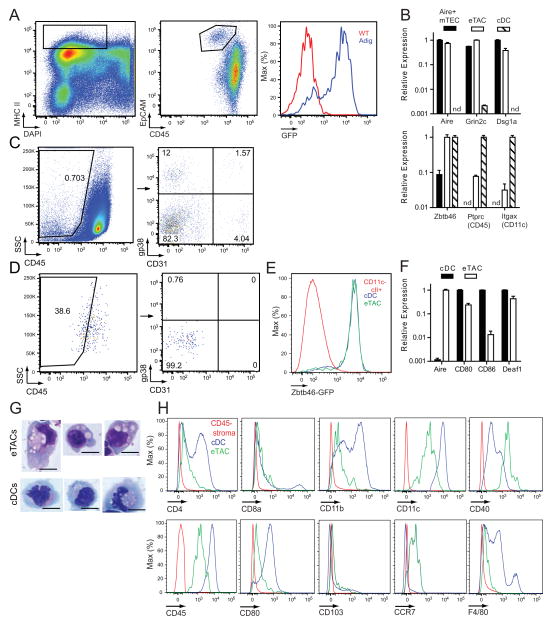
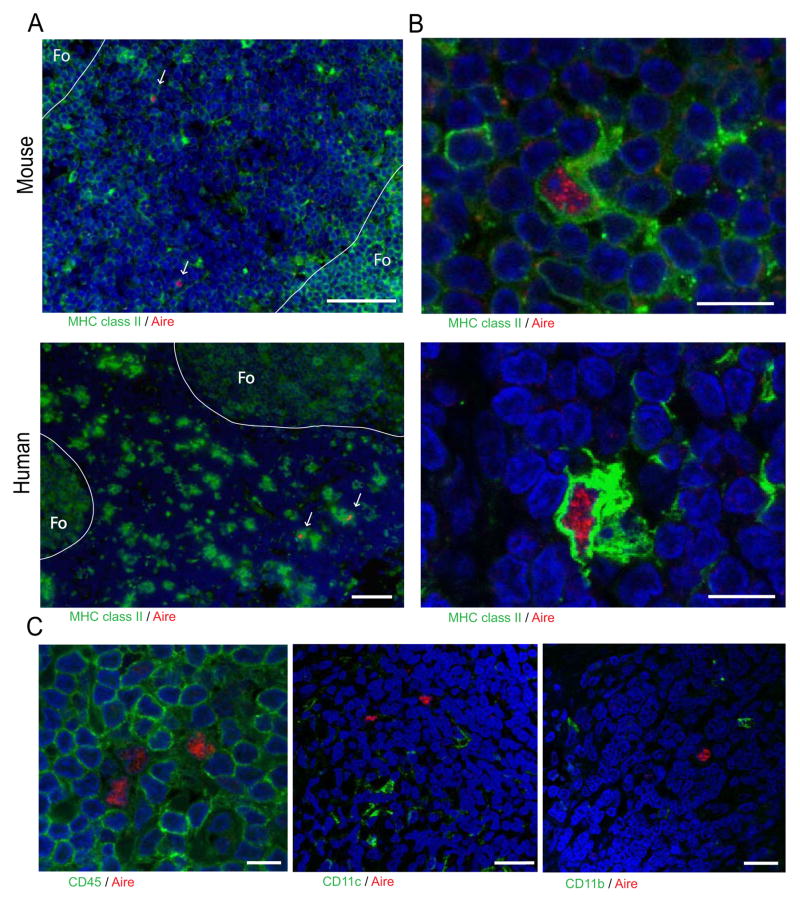
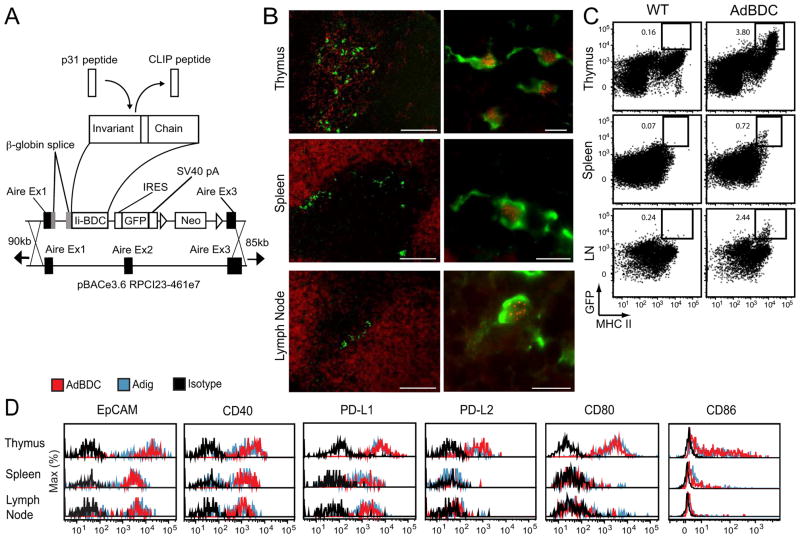
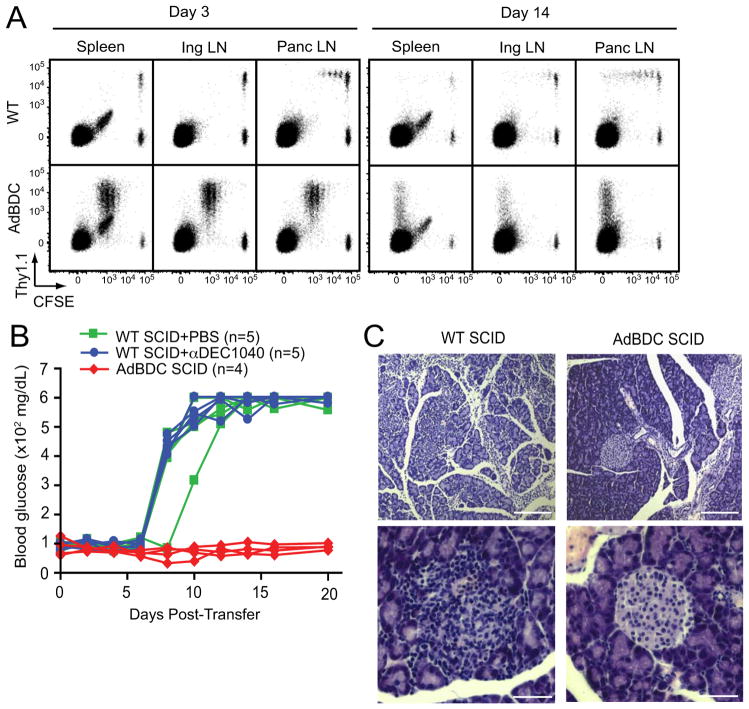
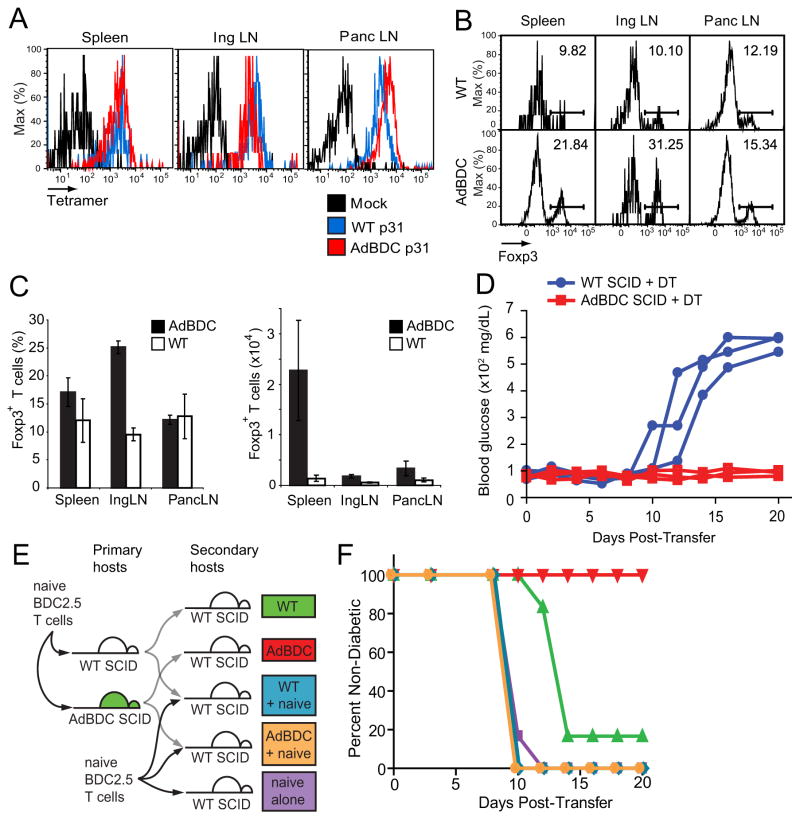
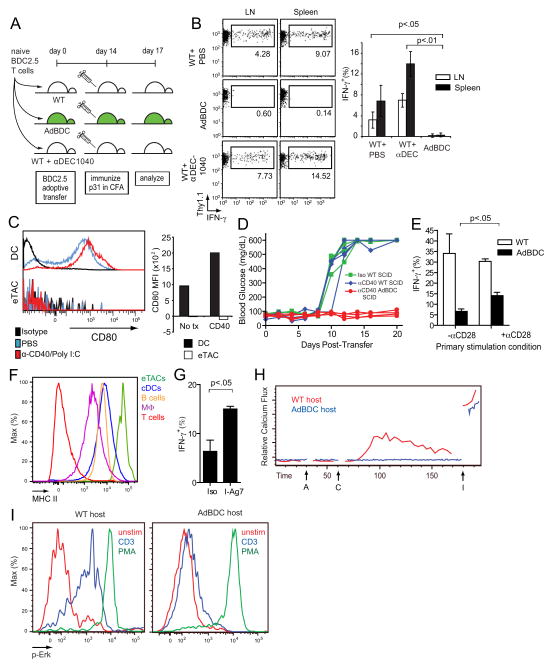
Comment in
-
A breath of fresh aire.Immunity. 2013 Sep 19;39(3):427-9. doi: 10.1016/j.immuni.2013.08.022. Immunity. 2013. PMID: 24054325
References
-
- Anderson MS, Venanzi ES, Chen Z, Berzins SP, Benoist C, Mathis D. The cellular mechanism of Aire control of T cell tolerance. Immunity. 2005;23:227–239. - PubMed
-
- Anderson MS, Venanzi ES, Klein L, Chen Z, Berzins SP, Turley SJ, von Boehmer H, Bronson R, Dierich A, Benoist C, et al. Projection of an immunological self shadow within the thymus by the aire protein. Science (New York, NY) 2002;298:1395–1401. - PubMed
-
- Bonifaz L, Bonnyay D, Mahnke K, Rivera M, Nussenzweig MC, Steinman RM. Efficient targeting of protein antigen to the dendritic cell receptor DEC-205 in the steady state leads to antigen presentation on major histocompatibility complex class I products and peripheral CD8+ T cell tolerance. The Journal of experimental medicine. 2002;196:1627–1638. - PMC - PubMed
-
- Chemnitz JM, Parry RV, Nichols KE, June CH, Riley JL. SHP-1 and SHP-2 associate with immunoreceptor tyrosine-based switch motif of programmed death 1 upon primary human T cell stimulation, but only receptor ligation prevents T cell activation. Journal of immunology (Baltimore, Md : 1950) 2004;173:945–954. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
