

Preconfigured, skewed distribution of firing rates in the hippocampus and entorhinal cortex
- PMID: 23994479
- PMCID: PMC3804159
- DOI: 10.1016/j.celrep.2013.07.039
Preconfigured, skewed distribution of firing rates in the hippocampus and entorhinal cortex
Abstract
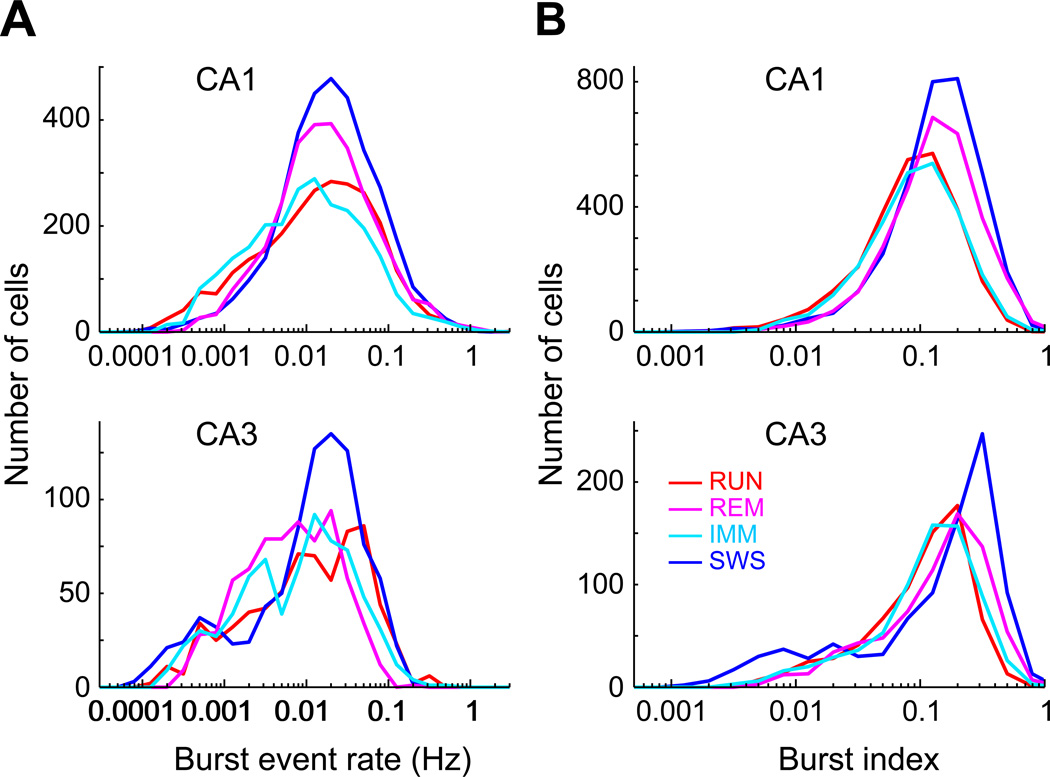
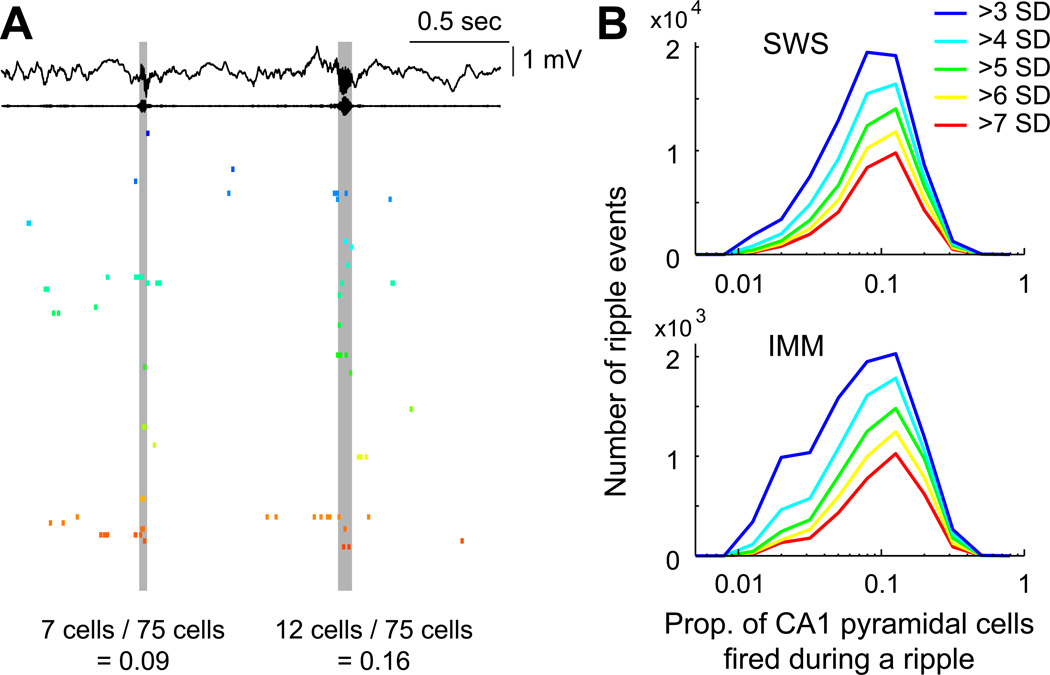
Despite the importance of the discharge frequency in neuronal communication, little is known about the firing-rate patterns of cortical populations. Using large-scale recordings from multiple layers of the entorhinal-hippocampal loop, we found that the firing rates of principal neurons showed a lognormal-like distribution in all brain states. Mean and peak rates within place fields of hippocampal neurons were also strongly skewed. Importantly, firing rates of the same neurons showed reliable correlations in different brain states and testing situations, as well as across familiar and novel environments. The fraction of neurons that participated in population oscillations displayed a lognormal pattern. Such skewed firing rates of individual neurons may be due to a skewed distribution of synaptic weights, which is supported by our observation of a lognormal distribution of the efficacy of spike transfer from principal neurons to interneurons. The persistent skewed distribution of firing rates implies that a preconfigured, highly active minority dominates information transmission in cortical networks.
Copyright © 2013 The Authors. Published by Elsevier Inc. All rights reserved.
Figures





References
-
- Barth AL, Poulet JF. Experimental evidence for sparse firing in the neocortex. Trends Neurosci. 2012;35:345–355. - PubMed
-
- Bartho P, Hirase H, Monconduit L, Zugaro M, Harris KD, Buzsaki G. Characterization of neocortical principal cells and interneurons by network interactions and extracellular features. J. Neurophysiol. 2004;92:600–608. - PubMed
-
- Buzsaki G, Horvath Z, Urioste R, Hetke J, Wise K. High-frequency network oscillation in the hippocampus. Science. 1992;256:1025–1027. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
