

Dietary tyrosine/phenylalanine depletion effects on behavioral and brain signatures of human motivational processing
- PMID: 23995581
- PMCID: PMC3895237
- DOI: 10.1038/npp.2013.232
Dietary tyrosine/phenylalanine depletion effects on behavioral and brain signatures of human motivational processing
Abstract
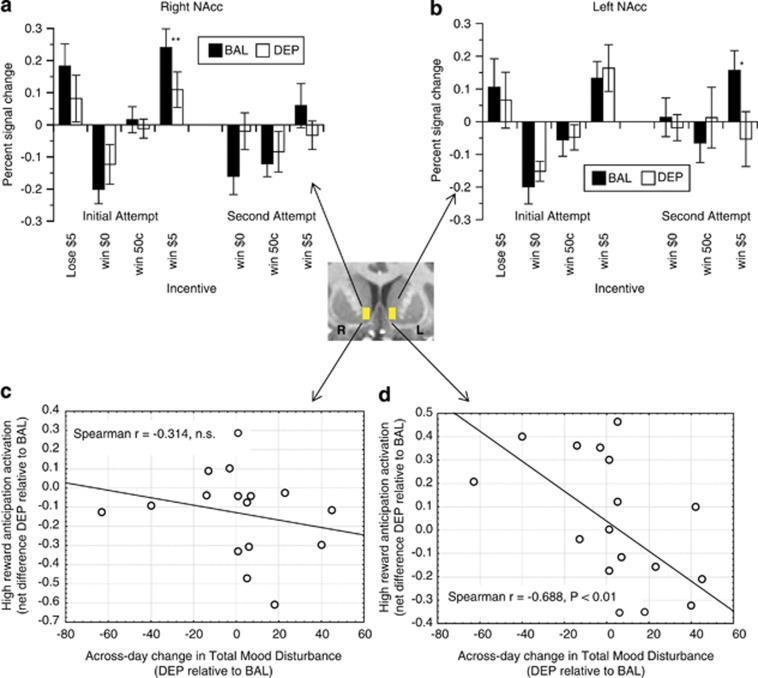
Dopamine (DA) neurotransmission is critical for motivational processing. We assessed whether disruption of DA synthesis in healthy controls using an amino-acid beverage devoid of catecholamine precursors (tyrosine-phenylalanine depletion (TPD)) would blunt recruitment of the nucleus accumbens (NAcc) by rewards. Sixteen controls ingested each of a tyr/phe-depleting beverage (DEP) or a tyr/phe-balanced (BAL) control beverage in two laboratory visits. Five hours after consumption of each drink, subjects underwent functional magnetic resonance imaging while they viewed anticipatory cues to respond to a target to either win money or avoid losing money. TPD did not exert main effects on mood or on task behavior, but affected brain activation. In right NAcc, TPD blunted activation by anticipation of high rewards. In left NAcc, recruitment anticipating high rewards was modulated by individual differences in mood change across the DEP drink day, where subjects whose mood worsened following TPD (relative to within-day mood change under BAL conditions) also showed lower activation under DEP conditions relative to BAL conditions. Exploratory analysis indicated that TPD qualitatively blunted the voxel-wise spatial extent of suprathreshold activation by reward anticipation. Finally, loss outcomes activated anterior insula under DEP conditions but not under BAL conditions. These data indicate that: (1) dietary depletion of catacholamine precursors will blunt dopaminergic mesolimbic activity, and (2) in controls, synthetic pathways of this neurocircuitry maintain sufficient buffering capacity to resist an effect on motivated behavior. Additional studies are needed to determine if clinical populations would show similar resistance to behavioral effects of TPD.
Figures




References
-
- Biggio G, Porceddu ML, Gessa GL. Decrease of homovanillic, dihydroxyphenylacetic acid and cyclic-adenosine-3′,5′-monophosphate content in the rat caudate nucleus induced by the acute administration of an aminoacid mixture lacking tyrosine and phenylalanine. J Neurochem. 1976;26:1253–1255. - PubMed
-
- Blum K, Braverman ER, Holder JM, Lubar JF, Monastra VJ, Miller D, et al. Reward deficiency syndrome: a biogenetic model for the diagnosis and treatment of impulsive, addictive, and compulsive behaviors. J Psychoactive Drugs. 2000;32 (Suppl i–iv:1–112. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
