

A nuclear F-actin scaffold stabilizes ribonucleoprotein droplets against gravity in large cells
- PMID: 23995731
- PMCID: PMC3789854
- DOI: 10.1038/ncb2830
A nuclear F-actin scaffold stabilizes ribonucleoprotein droplets against gravity in large cells
Abstract
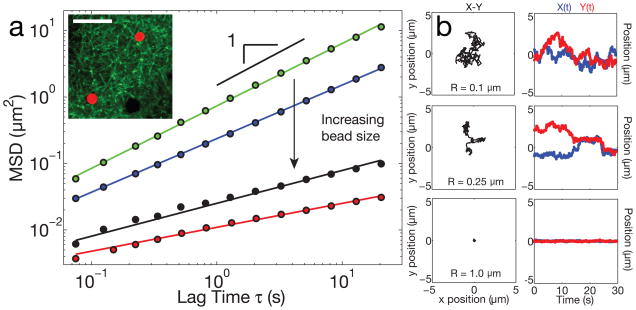
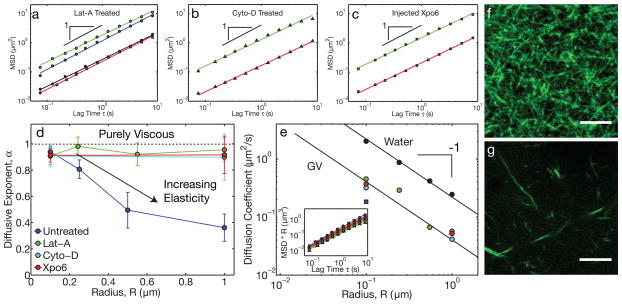
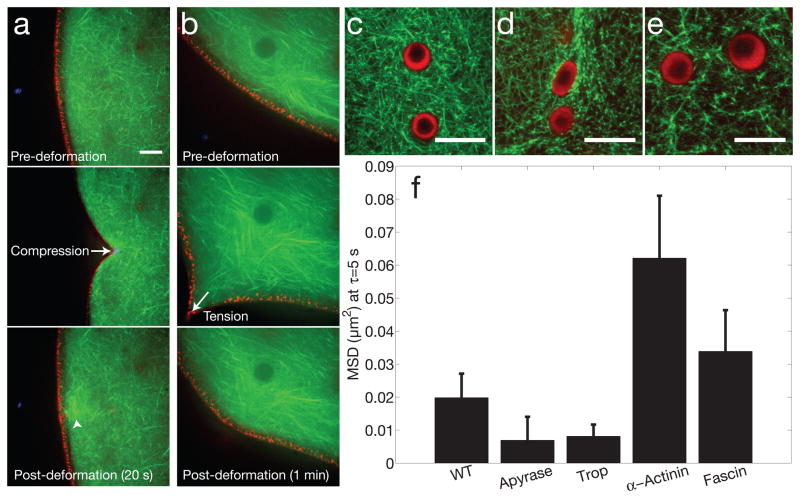
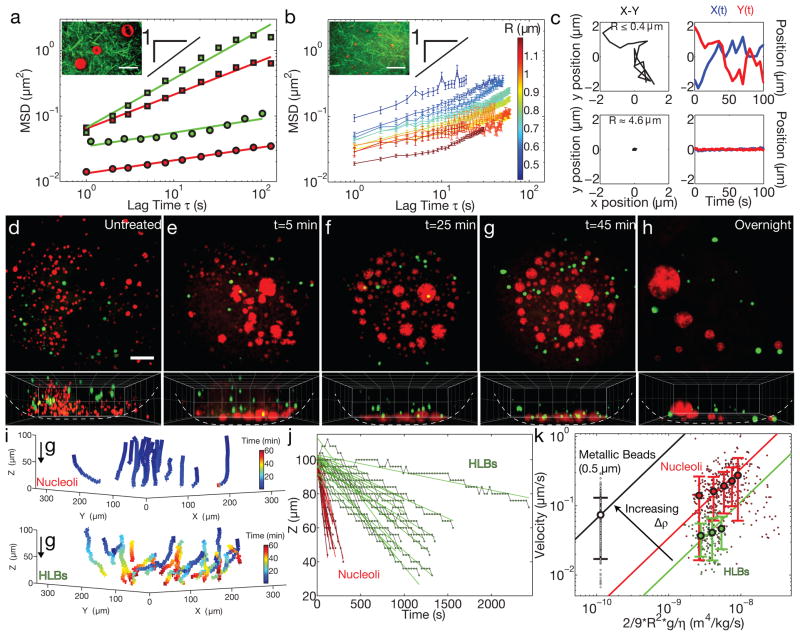
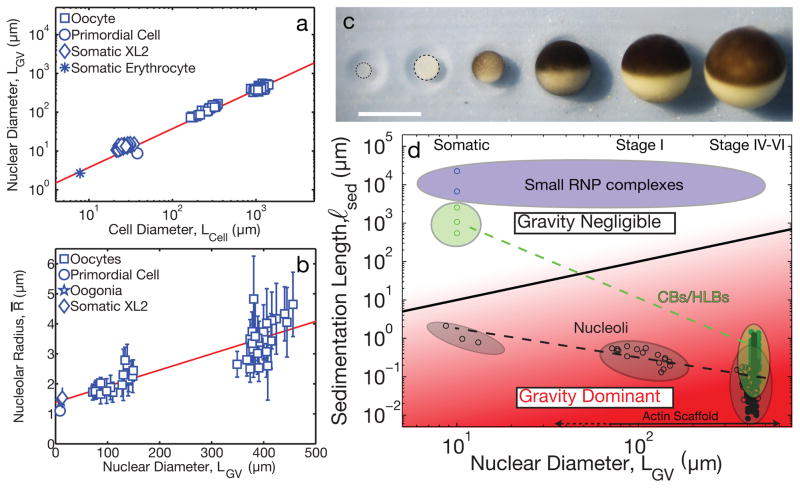
The size of a typical eukaryotic cell is of the order of ∼10 μm. However, some cell types grow to very large sizes, including oocytes (immature eggs) of organisms from humans to starfish. For example, oocytes of the frog Xenopus laevis grow to a diameter ≥1 mm. They have a correspondingly large nucleus (germinal vesicle) of ∼450 μm in diameter, which is similar to smaller somatic nuclei, but contains a significantly higher concentration of actin. The form and structure of this nuclear actin remain controversial, and its potential mechanical role within these large nuclei is unknown. Here, we use a microrheology and quantitative imaging approach to show that germinal vesicles contain an elastic F-actin scaffold that mechanically stabilizes these large nuclei against gravitational forces, which are usually considered negligible within cells. We find that on actin disruption, ribonucleoprotein droplets, including nucleoli and histone locus bodies, undergo gravitational sedimentation and fusion. We develop a model that reveals how gravity becomes an increasingly potent force as cells and their nuclei grow larger than ∼10 μm, explaining the requirement for a stabilizing nuclear F-actin scaffold in large Xenopus oocytes. All life forms are subject to gravity, and our results may have broad implications for cell growth and size control.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
-
- Lammerding J, Dahl KN, Discher DE, Kamm RD. In: Methods in Cell Biology. Yu-Li Wang, Discher Dennis E., editors. Vol. 83. Academic Press; 2007. pp. 269–294. - PubMed
-
- Rando OJ, Zhao K, Crabtree GR. Searching for a function for nuclear actin. Trends Cell Biol. 2000;10:92–97. S0962-8924(99)01713-4 [pii] - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
