

The structural basis for phospholamban inhibition of the calcium pump in sarcoplasmic reticulum
- PMID: 23996003
- PMCID: PMC3798486
- DOI: 10.1074/jbc.M113.501585
The structural basis for phospholamban inhibition of the calcium pump in sarcoplasmic reticulum
Abstract
P-type ATPases are a large family of enzymes that actively transport ions across biological membranes by interconverting between high (E1) and low (E2) ion-affinity states; these transmembrane transporters carry out critical processes in nearly all forms of life. In striated muscle, the archetype P-type ATPase, SERCA (sarco(endo)plasmic reticulum Ca(2+)-ATPase), pumps contractile-dependent Ca(2+) ions into the lumen of sarcoplasmic reticulum, which initiates myocyte relaxation and refills the sarcoplasmic reticulum in preparation for the next contraction. In cardiac muscle, SERCA is regulated by phospholamban (PLB), a small inhibitory phosphoprotein that decreases the Ca(2+) affinity of SERCA and attenuates contractile strength. cAMP-dependent phosphorylation of PLB reverses Ca(2+)-ATPase inhibition with powerful contractile effects. Here we present the long sought crystal structure of the PLB-SERCA complex at 2.8-Å resolution. The structure was solved in the absence of Ca(2+) in a novel detergent system employing alkyl mannosides. The structure shows PLB bound to a previously undescribed conformation of SERCA in which the Ca(2+) binding sites are collapsed and devoid of divalent cations (E2-PLB). This new structure represents one of the key unsolved conformational states of SERCA and provides a structural explanation for how dephosphorylated PLB decreases Ca(2+) affinity and depresses cardiac contractility.
Keywords: Calcium ATPase; Calcium Transport; Crystal Structure; Phospholamban; Protein Cross-linking; Protein Crystallization; SERCA; Sarcolipin; Sarcoplasmic Reticulum (SR).
Figures
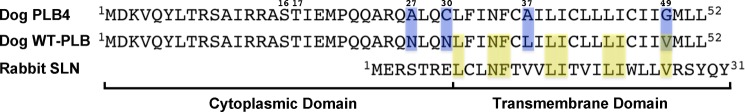


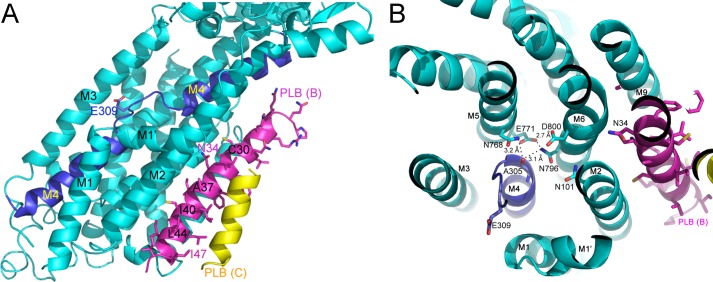
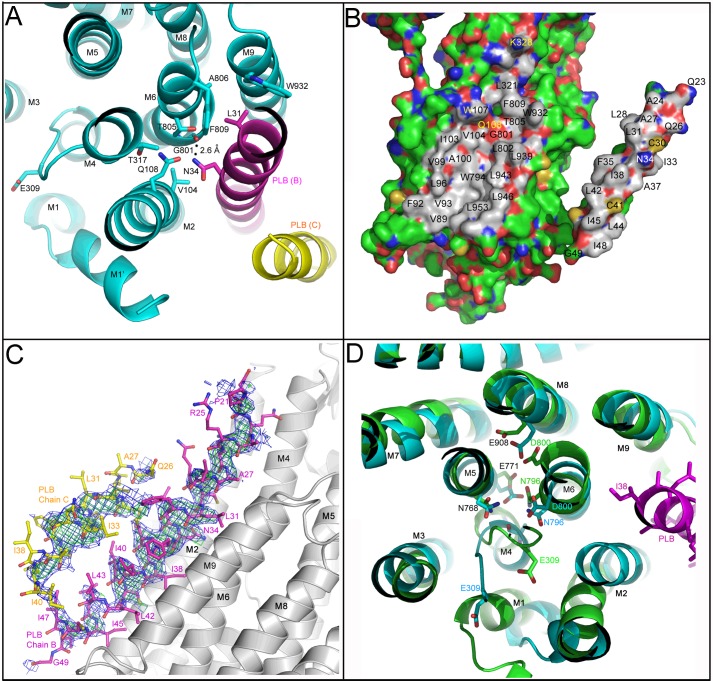
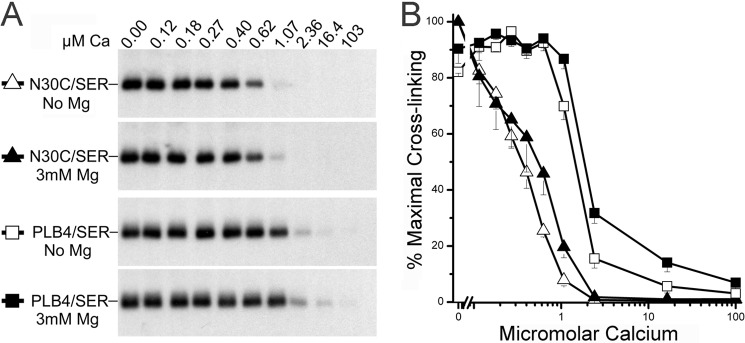
References
-
- Lindemann J. P., Jones L. R., Hathaway D. R., Henry B. G., Watanabe A. M. (1983) β-Adrenergic stimulation of phospholamban phosphorylation and Ca2+-ATPase activity in guinea pig ventricles. J. Biol. Chem. 258, 464–471 - PubMed
-
- Simmerman H. K., Jones L. R. (1998) Phospholamban. Protein structure, mechanism of action, and role in cardiac function. Physiol. Rev. 78, 921–947 - PubMed
-
- Young H. S., Stokes D. L. (2004) The mechanics of calcium transport. J. Membr. Biol. 198, 55–63 - PubMed
-
- Wegener A. D., Simmerman H. K., Lindemann J. P., Jones L. R. (1989) Phospholamban phosphorylation in intact ventricles. Phosphorylation of serine 16 and threonine 17 in response to β-adrenergic stimulation. J. Biol. Chem. 264, 11468–11474 - PubMed
-
- Jones L. R., Cornea R. L., Chen Z. (2002) Close proximity between residue 30 of phospholamban and cysteine 318 of the cardiac Ca2+ pump revealed by intermolecular thiol cross-linking. J. Biol. Chem. 277, 28319–28329 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
