

Engineering modular 'ON' RNA switches using biological components
- PMID: 23999097
- PMCID: PMC3905868
- DOI: 10.1093/nar/gkt787
Engineering modular 'ON' RNA switches using biological components
Abstract
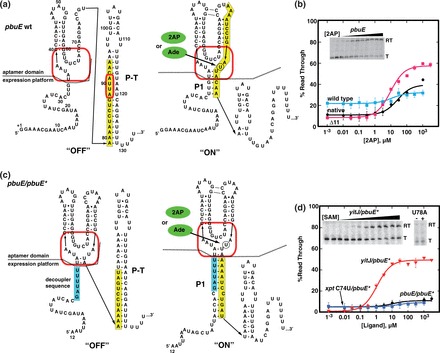
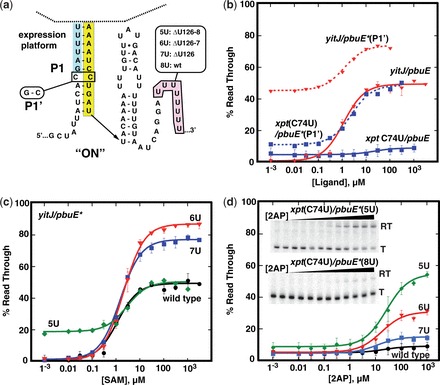
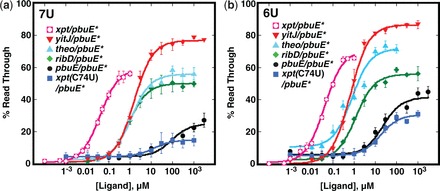
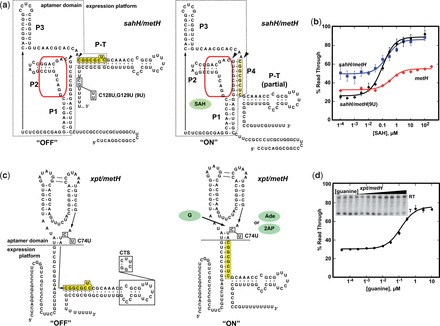
Riboswitches are cis-acting regulatory elements broadly distributed in bacterial mRNAs that control a wide range of critical metabolic activities. Expression is governed by two distinct domains within the mRNA leader: a sensory 'aptamer domain' and a regulatory 'expression platform'. Riboswitches have also received considerable attention as important tools in synthetic biology because of their conceptually simple structure and the ability to obtain aptamers that bind almost any conceivable small molecule using in vitro selection (referred to as SELEX). In the design of artificial riboswitches, a significant hurdle has been to couple the two domains enabling their efficient communication. We previously demonstrated that biological transcriptional 'OFF' expression platforms are easily coupled to diverse aptamers, both biological and SELEX-derived, using simple design rules. Here, we present two modular transcriptional 'ON' riboswitch expression platforms that are also capable of hosting foreign aptamers. We demonstrate that these biological parts can be used to facilely generate artificial chimeric riboswitches capable of robustly regulating transcription both in vitro and in vivo. We expect that these modular expression platforms will be of great utility for various synthetic biological applications that use RNA-based biosensors.
Figures
Similar articles
-
Design of modular "plug-and-play" expression platforms derived from natural riboswitches for engineering novel genetically encodable RNA regulatory devices.Methods Enzymol. 2015;550:41-71. doi: 10.1016/bs.mie.2014.10.031. Epub 2014 Dec 19. Methods Enzymol. 2015. PMID: 25605380
-
RNA aptamers as genetic control devices: the potential of riboswitches as synthetic elements for regulating gene expression.Biotechnol J. 2015 Feb;10(2):246-57. doi: 10.1002/biot.201300498. Biotechnol J. 2015. PMID: 25676052 Review.
-
Engineering and In Vivo Applications of Riboswitches.Annu Rev Biochem. 2017 Jun 20;86:515-539. doi: 10.1146/annurev-biochem-060815-014628. Epub 2017 Mar 30. Annu Rev Biochem. 2017. PMID: 28375743 Review.
-
Modularity of select riboswitch expression platforms enables facile engineering of novel genetic regulatory devices.ACS Synth Biol. 2013 Aug 16;2(8):463-72. doi: 10.1021/sb4000096. Epub 2013 Mar 28. ACS Synth Biol. 2013. PMID: 23654267 Free PMC article.
-
Development of Novel Riboswitches for Synthetic Biology in the Green Alga Chlamydomonas.ACS Synth Biol. 2020 Jun 19;9(6):1406-1417. doi: 10.1021/acssynbio.0c00082. Epub 2020 Jun 4. ACS Synth Biol. 2020. PMID: 32496044 Free PMC article.
Cited by
-
Structure-guided mutational analysis of gene regulation by the Bacillus subtilis pbuE adenine-responsive riboswitch in a cellular context.J Biol Chem. 2015 Feb 13;290(7):4464-75. doi: 10.1074/jbc.M114.613497. Epub 2014 Dec 30. J Biol Chem. 2015. PMID: 25550163 Free PMC article.
-
Regulatory RNAs in Bacillus subtilis: A review on regulatory mechanism and applications in synthetic biology.Synth Syst Biotechnol. 2024 Feb 10;9(2):223-233. doi: 10.1016/j.synbio.2024.01.013. eCollection 2024 Jun. Synth Syst Biotechnol. 2024. PMID: 38385150 Free PMC article. Review.
-
Small synthetic molecule-stabilized RNA pseudoknot as an activator for -1 ribosomal frameshifting.Nucleic Acids Res. 2018 Sep 19;46(16):8079-8089. doi: 10.1093/nar/gky689. Nucleic Acids Res. 2018. PMID: 30085309 Free PMC article.
-
Evaluating riboswitch optimality.Methods Enzymol. 2019;623:417-450. doi: 10.1016/bs.mie.2019.05.028. Epub 2019 Jun 18. Methods Enzymol. 2019. PMID: 31239056 Free PMC article.
-
Elimination of editing plasmid mediated by theophylline riboswitch in Zymomonas mobilis.Appl Microbiol Biotechnol. 2023 Dec;107(23):7151-7163. doi: 10.1007/s00253-023-12783-y. Epub 2023 Sep 20. Appl Microbiol Biotechnol. 2023. PMID: 37728624
References
-
- Keasling JD. Synthetic biology and the development of tools for metabolic engineering. Metab. Eng. 2012;14:189–195. - PubMed
-
- Benenson Y. Synthetic biology with RNA: progress report. Curr. Opin. Chem. Biol. 2012;16:278–284. - PubMed
-
- Wieland M, Fussenegger M. Ligand-dependent regulatory RNA parts for synthetic biology in eukaryotes. Curr. Opin. Chem. Biol. 2010;21:760–765. - PubMed
-
- Isaacs FJ, Dwyer DJ, Collins JJ. RNA synthetic biology. Nat. Biotechnol. 2006;24:545–554. - PubMed
-
- Wittmann A, Suess B. Engineered riboswitches: expanding researchers' toolbox with synthetic RNA regulators. FEBS Lett. 2012;586:2076–2083. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
