

Immunodominance and functional alterations of tumor-associated antigen-specific CD8+ T-cell responses in hepatocellular carcinoma
- PMID: 24002931
- PMCID: PMC4139003
- DOI: 10.1002/hep.26731
Immunodominance and functional alterations of tumor-associated antigen-specific CD8+ T-cell responses in hepatocellular carcinoma
Abstract
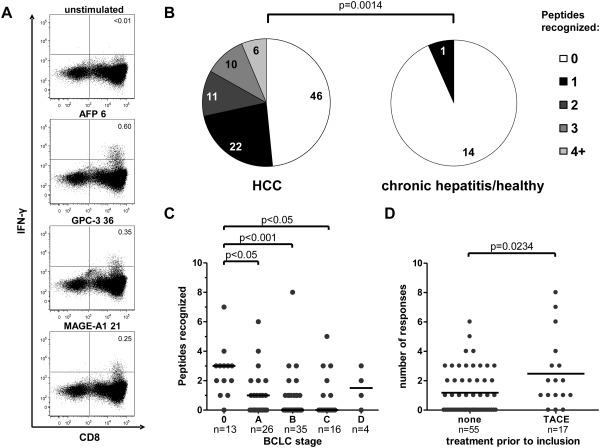
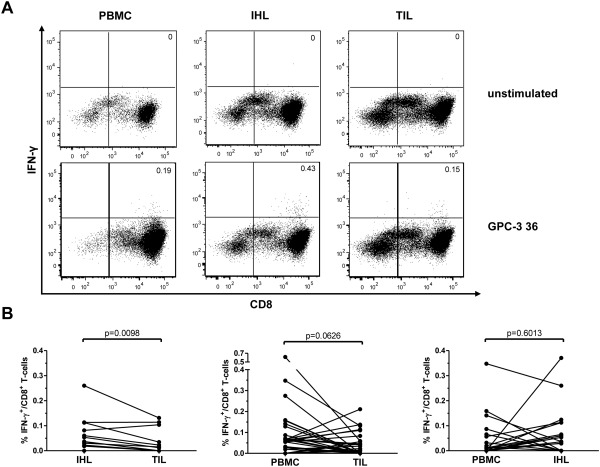
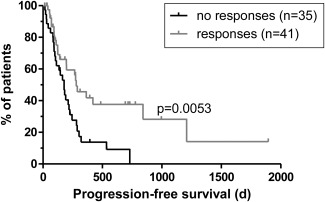
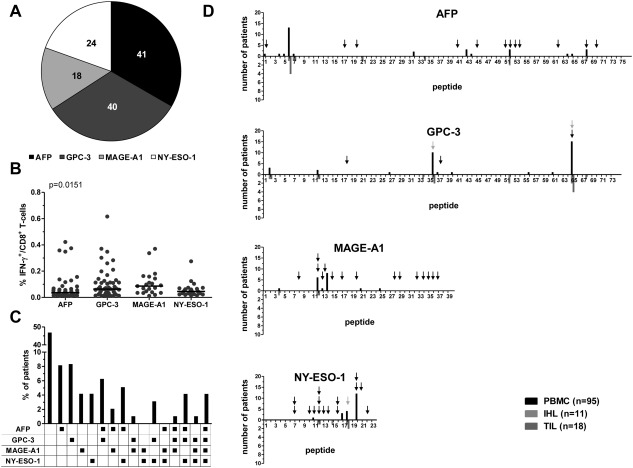
Hepatocellular carcinoma (HCC) is the fifth most common malignancy worldwide with a poor prognosis and limited therapeutic options. To aid the development of novel immunological interventions, we studied the breadth, frequency, and tumor-infiltration of naturally occurring CD8(+) T-cell responses targeting several tumor-associated antigens (TAA). We used overlapping peptides spanning the entire alpha-fetoprotein (AFP), glypican-3 (GPC-3), melanoma-associated gene-A1 (MAGE-A1) and New York-esophageal squamous cell carcinoma-1 (NY-ESO-1) proteins and major-histocompatibility-complex-class-I-tetramers specific for epitopes of MAGE-A1 and NY-ESO-1 to analyze TAA-specific CD8(+) T-cell responses in a large cohort of HCC patients. After nonspecific expansion in vitro, we detected interferon-γ (IFN-γ)-producing CD8(+) T cells specific for all four TAA in the periphery as well as in liver and tumor tissue. These CD8(+) T-cell responses displayed clear immunodominance patterns within each TAA, but no consistent hierarchy was observed between different TAA. Importantly, the response breadth was highest in early-stage HCC and associated with patient survival. After antigen-specific expansion, TAA-specific CD8(+) T cells were detectable by tetramer staining but impaired in their ability to produce IFN-γ. Furthermore, regulatory T cells (Treg) were increased in HCC lesions. Depletion of Treg from cultures improved TAA-specific CD8(+) T-cell proliferation but did not restore IFN-γ-production.
Conclusion: Naturally occurring TAA-specific CD8(+) T-cell responses are present in patients with HCC and therefore constitute part of the normal T-cell repertoire. Moreover, the presence of these responses correlates with patient survival. However, the observation of impaired IFN-γ production suggests that the efficacy of such responses is functionally limited. These findings support the development of strategies that aim to enhance the total TAA-specific CD8(+) T-cell response by therapeutic boosting and/or specificity diversification. However, further research will be required to help unlock the full potential of TAA-specific CD8(+) T-cell responses.
© 2014 The Authors. Hepatology published by Wiley on behalf of the American Association for the Study of Liver Diseases.
Conflict of interest statement
Potential conflict of interest: Nothing to report.
Figures
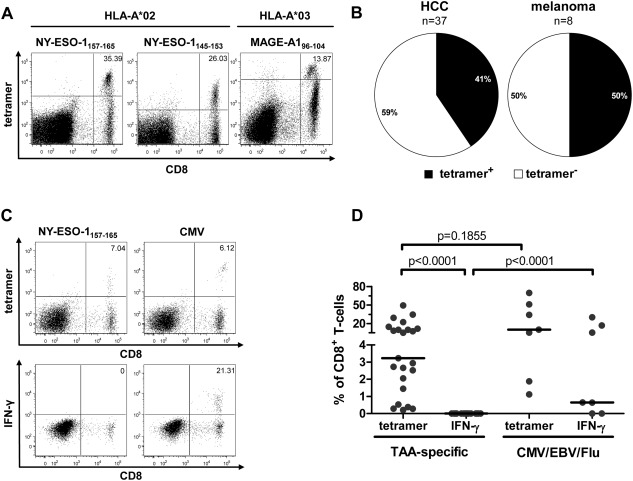
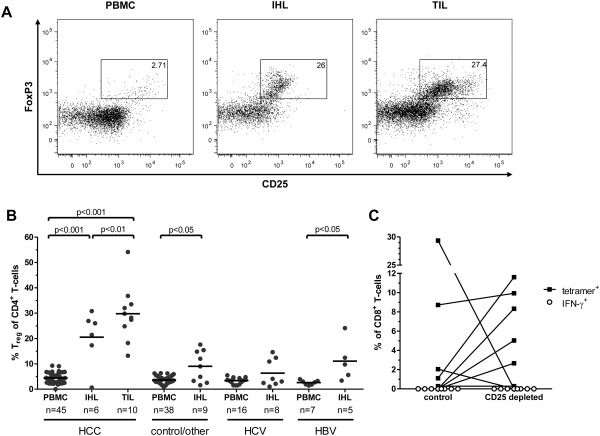
Comment in
-
Antitumor CD8+ T cells in hepatocellular carcinoma: present but exhausted.Hepatology. 2014 Apr;59(4):1232-4. doi: 10.1002/hep.26779. Epub 2014 Feb 18. Hepatology. 2014. PMID: 24123091 No abstract available.
References
-
- El‐Serag HB. Hepatocellular carcinoma. N Engl J Med 2011;365:1118–1127 - PubMed
-
- Yoong KF, McNab G, Hubscher SG, Adams DH. Vascular adhesion protein‐1 and ICAM‐1 support the adhesion of tumor‐infiltrating lymphocytes to tumor endothelium in human hepatocellular carcinoma. J Immunol 1998;160:3978–3988 - PubMed
-
- Wada Y, Nakashima O, Kutami R, Yamamoto O, Kojiro M. Clinicopathological study on hepatocellular carcinoma with lymphocytic infiltration. Hepatology 1998;27:407–414 - PubMed
-
- Takayama T, Sekine T, Makuuchi M, Yamasaki S, Kosuge T, Yamamoto J, et al. Adoptive immunotherapy to lower postsurgical recurrence rates of hepatocellular carcinoma: a randomised trial. Lancet 2000;356:802–807 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials