

Endogenous interleukin-1β in neuropathic rats enhances glutamate release from the primary afferents in the spinal dorsal horn through coupling with presynaptic N-methyl-D-aspartic acid receptors
- PMID: 24003233
- PMCID: PMC3798525
- DOI: 10.1074/jbc.M113.495465
Endogenous interleukin-1β in neuropathic rats enhances glutamate release from the primary afferents in the spinal dorsal horn through coupling with presynaptic N-methyl-D-aspartic acid receptors
Abstract
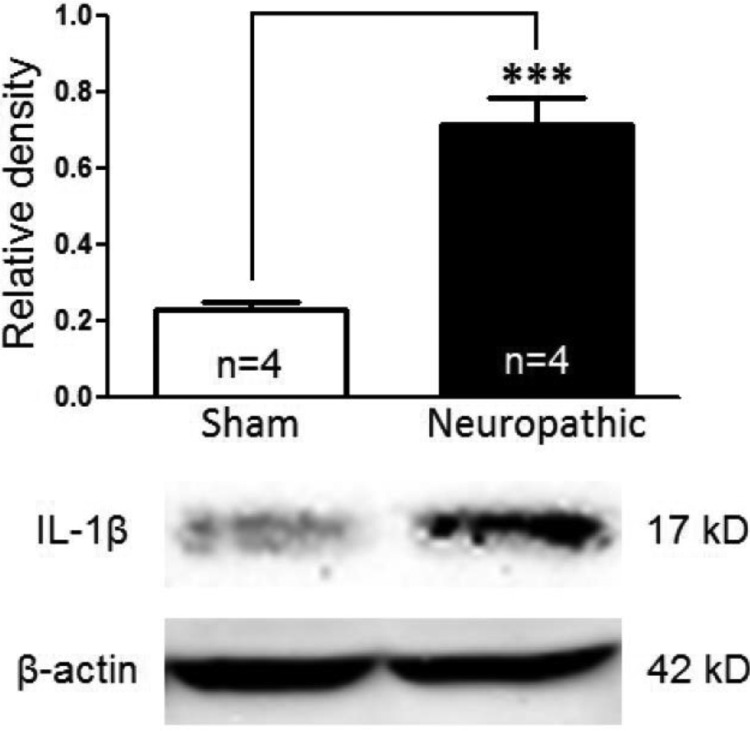
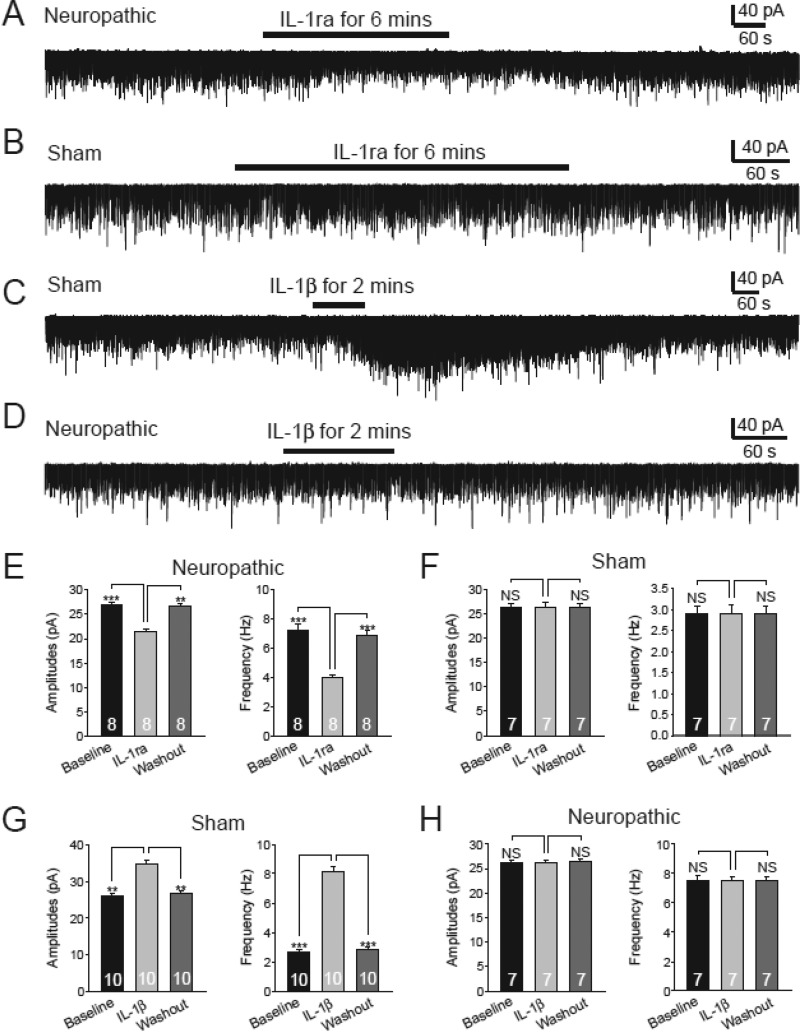
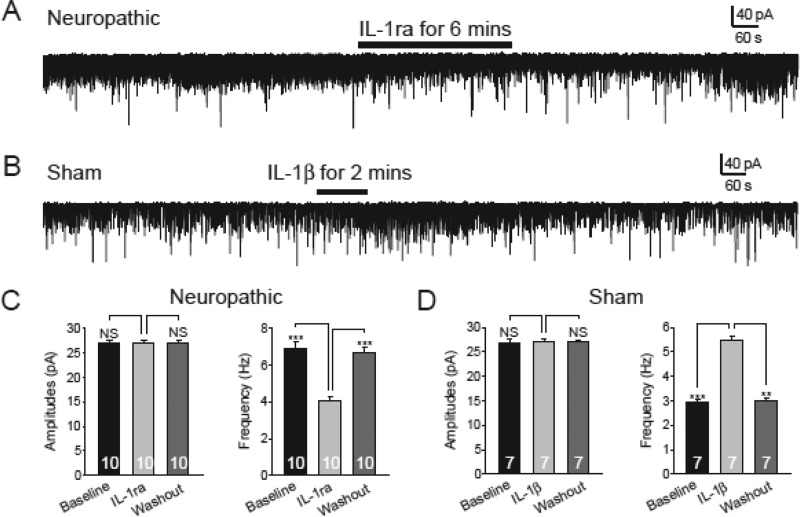
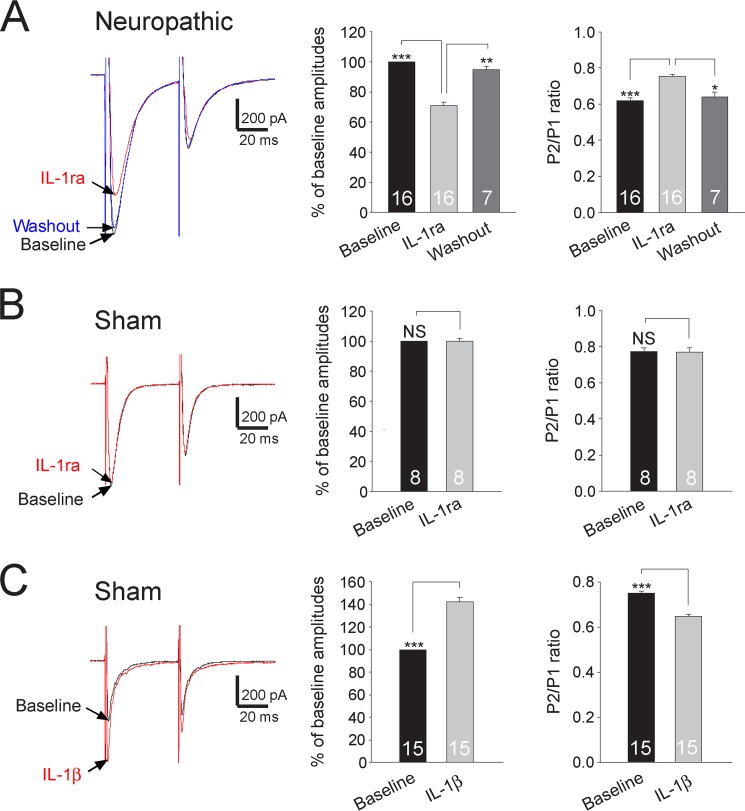
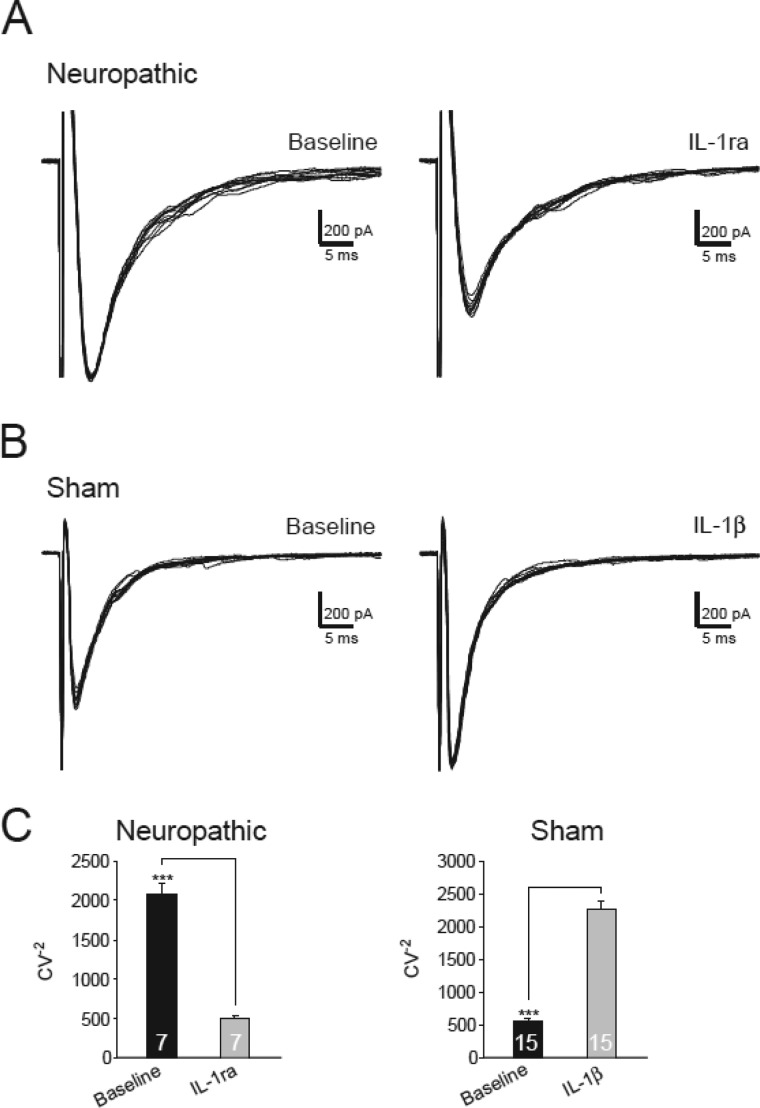
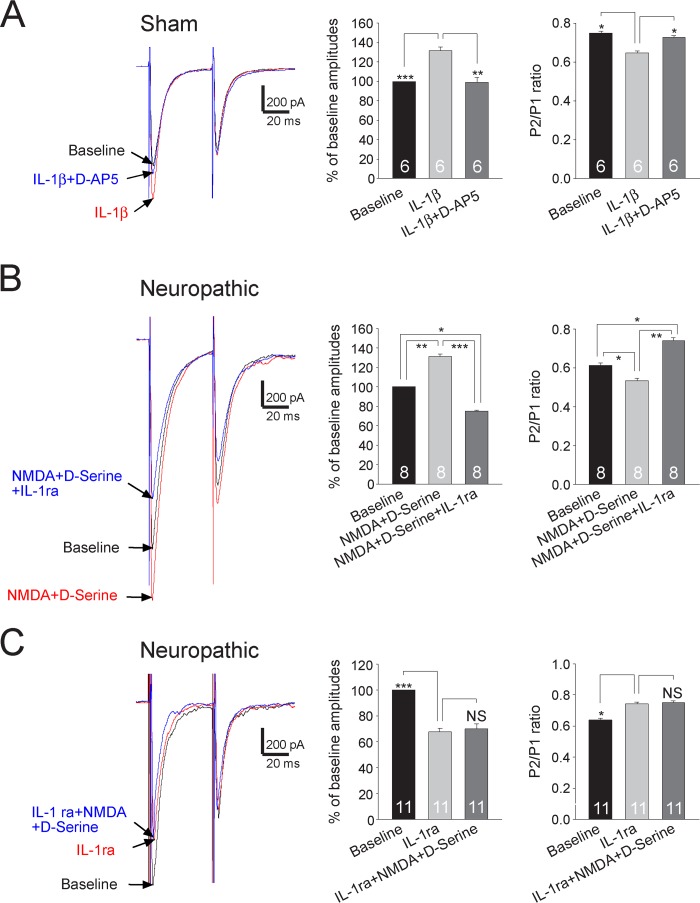
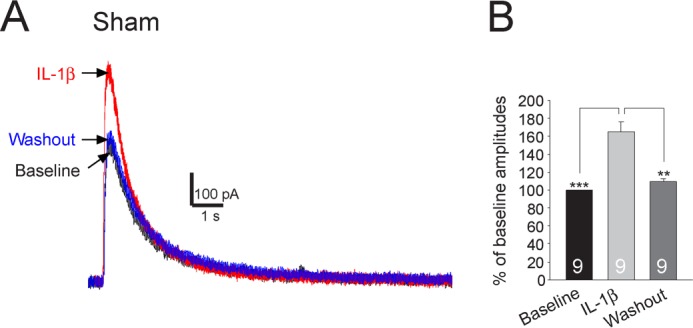
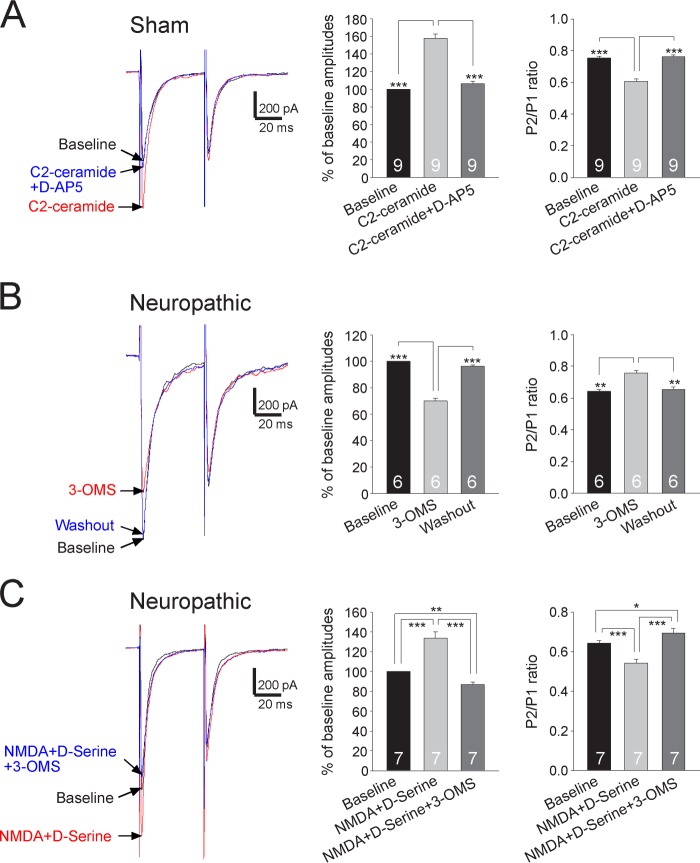
Excessive activation of glutamate receptors and overproduction of proinflammatory cytokines, including interleukin-1β (IL-1β) in the spinal dorsal horn, are key mechanisms underlying the development and maintenance of neuropathic pain. In this study, we investigated the mechanisms by which endogenous IL-1β alters glutamatergic synaptic transmission in the spinal dorsal horn in rats with neuropathic pain induced by ligation of the L5 spinal nerve. We demonstrated that endogenous IL-1β in neuropathic rats enhances glutamate release from the primary afferent terminals and non-NMDA glutamate receptor activities in postsynaptic neurons in the spinal dorsal horn. Myeloid differentiation primary response protein 88 (MyD88) is a mediator used by IL-1β to enhance non-NMDA glutamate receptor activities in postsynaptic neurons in the spinal dorsal horn. Presynaptic NMDA receptors are effector receptors used by the endogenous IL-1β to enhance glutamate release from the primary afferents in neuropathic rats. This is further supported by the fact that NMDA currents recorded from small neurons in the dorsal root ganglion of normal rats are potentiated by exogenous IL-1β. Furthermore, we provided evidence that functional coupling between IL-1β receptors and presynaptic NMDA receptors at the primary afferent terminals is mediated by the neutral sphingomyelinase/ceramide signaling pathway. Hence, functional coupling between IL-1β receptors and presynaptic NMDA receptors at the primary afferent terminals is a crucial mechanism leading to enhanced glutamate release and activation of non-NMDA receptors in the spinal dorsal horn neurons in neuropathic pain conditions. Interruption of such functional coupling could be an effective approach for the treatment of neuropathic pain.
Keywords: Cell Signaling; Glia; Glutamate Receptors Ionotropic (AMPA, NMDA); Pain; Patch Clamp.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
