

Evidence for a diffusible factor that induces susceptibility in the Colletotrichum-maize disease interaction
- PMID: 24003973
- PMCID: PMC6638722
- DOI: 10.1111/mpp.12069
Evidence for a diffusible factor that induces susceptibility in the Colletotrichum-maize disease interaction
Abstract
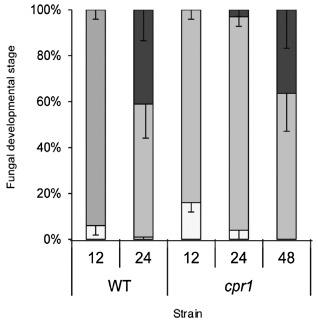
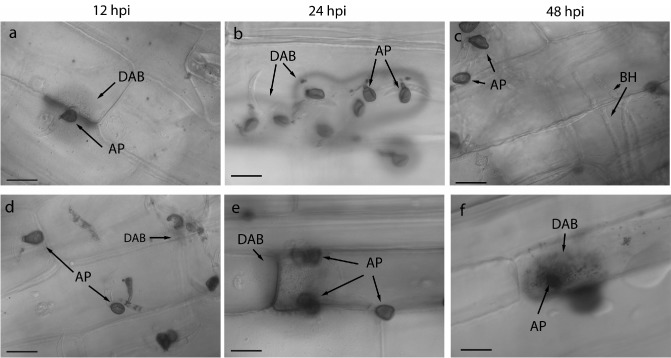
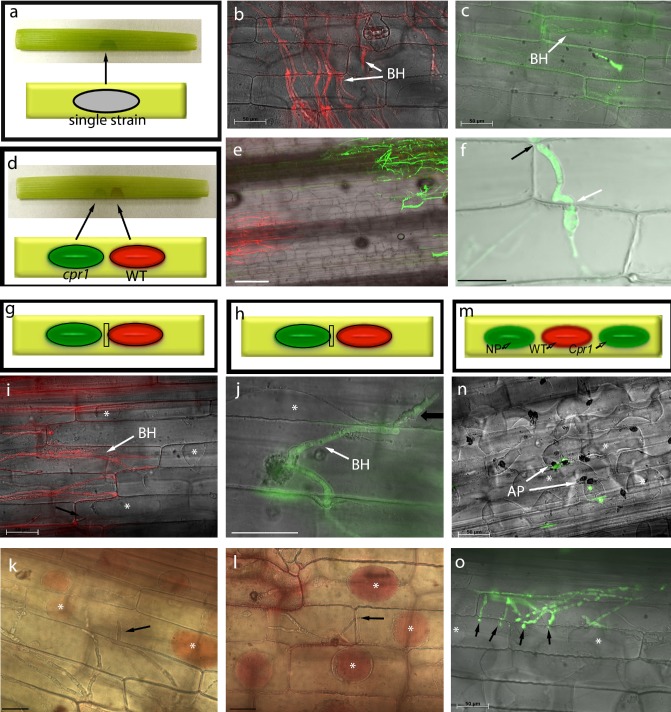
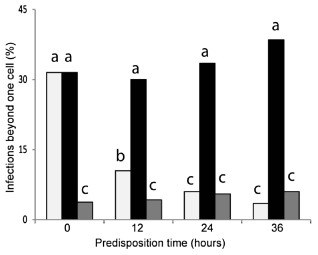
Colletotrichum graminicola, the causal agent of maize anthracnose, is a hemibiotrophic fungus that initially infects living host cells via primary hyphae surrounded by a membrane. A nonpathogenic mutant disrupted in a gene encoding a component of the signal peptidase complex, and believed to be deficient in protein processing and secretion, regained pathogenicity when it was inoculated onto maize leaf sheaths close to the wild-type fungus. Evidence is presented suggesting that the wild-type produces a diffusible factor(s) that induces the localized susceptibility of host cells at the borders of expanding colonies, causing them to become receptive to biotrophic invasion. The induced susceptibility effect is limited to a distance of approximately eight cells from the edge of the wild-type colony, is dosage dependent and is specific to C. graminicola.
© 2013 BSPP AND JOHN WILEY & SONS LTD.
Figures

References
-
- Amselem, J. , Cuomo, C.A. , van Kan, J.A.L. , Viaud, M. , Benito, E.P. , Couloux, A. , Coutinho, P.M. , de Vriesm, R.P. , Dyer, P.S. , Fillinger, S. , Fournier, E. , Gout, L. , Hahn, M. , Kohn, L. , Lapalu, N. , Plummer, K.M. , Pradier, J.M. , Quévillon, E. , Sharon, A. , Simon, A. , ten Have, A. , Tudzynski, B. , Tudzynski, P. , Wincker, P. , Andrew, M. , Anthouard, V. , Beever, R.E. , Beffa, R. , Benoit, I. , Bouzid, O. , Brault, B. , Chen, Z. , Choquer, M. , Collémare, J. , Cotton, P. , Danchin, E.G. , Da Silva, C. , Gautier, A. , Giraud, C. , Giraud, T. , Gonzalez, C. , Grossetete, S. , Güldener, U. , Henrissat, B. , Howlett, B.J. , Kodira, C. , Kretschmer, M. , Lappartient, A. , Leroch, M. , Levis, C. , Mauceli, E. , Neuvéglise, C. , Oeser, B. , Pearson, M. , Poulain, J. , Poussereau, N. , Quesneville, H. , Rascle, C. , Schumacher, J. , Ségurens, B. , Sexton, A. , Silva, E. , Sirven, C. , Soanes, D.M. , Talbot, N.J. , Templeton, M. , Yandava, C. , Yarden, O. , Zeng, Q. , Rollins, J.A. , Lebrun, M.H. and Dickman, M. (2011) Genomic analysis of the necrotrophic fungal pathogens Sclerotinia sclerotiorum and Botrytis cinerea . PLoS Genet. 7, e1002230. - PMC - PubMed
-
- Andrie, R.M. , Martinez, J.P. and Ciuffetti, L.M. (2005) Development of ToxA and ToxB promoter‐driven fluorescent protein expression vectors for use in filamentous ascomycetes. Mycologia, 97, 1152–1161. - PubMed
-
- Bell, A. and Presley, J. (1969) Heat‐inhibited or heat‐killed conidia of Verticillium albo‐atrum induce disease resistance and phytoalexin synthesis in cotton. Phytopathology, 59, 1147–1151.
-
- Benito, E.P. , ten Have, A. , van't Klooster, J.W. and van Kan, J.A. (1998) Fungal and plant gene expression during synchronized infection of tomato leaves by Botrytis cinerea . Eur. J. Plant Pathol. 104, 207–220.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
