

Erythropoietin is a JAK2 and ERK1/2 effector that can promote renal tumor cell proliferation under hypoxic conditions
- PMID: 24004818
- PMCID: PMC3844377
- DOI: 10.1186/1756-8722-6-65
Erythropoietin is a JAK2 and ERK1/2 effector that can promote renal tumor cell proliferation under hypoxic conditions
Abstract
Background: Erythropoietin (EPO) provides an alternative to transfusion for increasing red blood cell mass and treating anemia in cancer patients. However, recent studies have reported increased adverse events and/or reduced survival in patients receiving both EPO and chemotherapy, potentially related to EPO-induced cancer progression. Additional preclinical studies that elucidate the possible mechanism underlying EPO cellular growth stimulation are needed.
Methods: Using commercial tissue microarray (TMA) of a variety of cancers and benign tissues, EPO and EPO receptor immunohistochemical staining was performed. Furthermore using a panel of human renal cells (Caki-1, 786-O, 769-P, RPTEC), in vitro and in vivo experiments were performed with the addition of EPO in normoxic and hypoxic states to note phenotypic and genotypic changes.
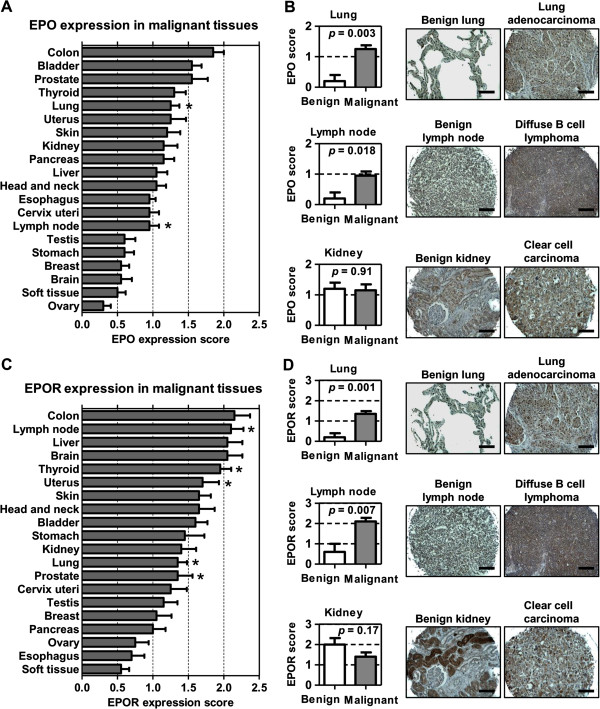
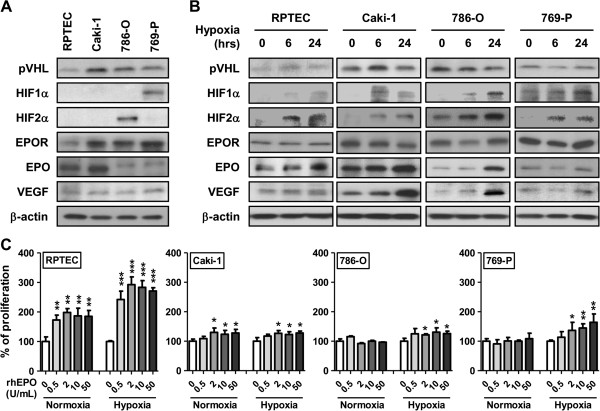
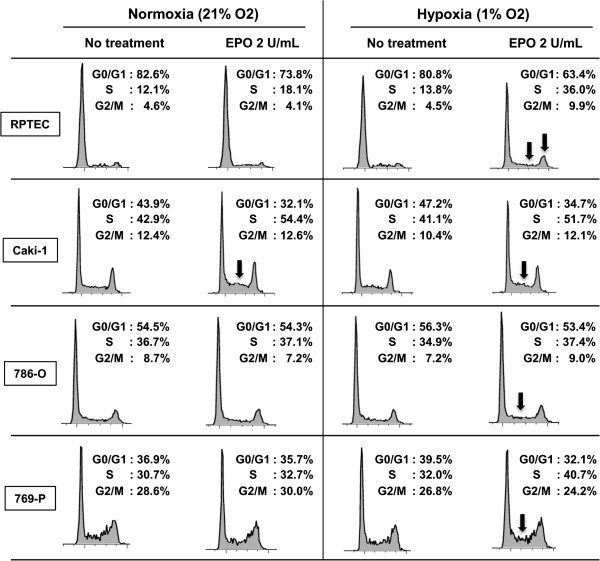
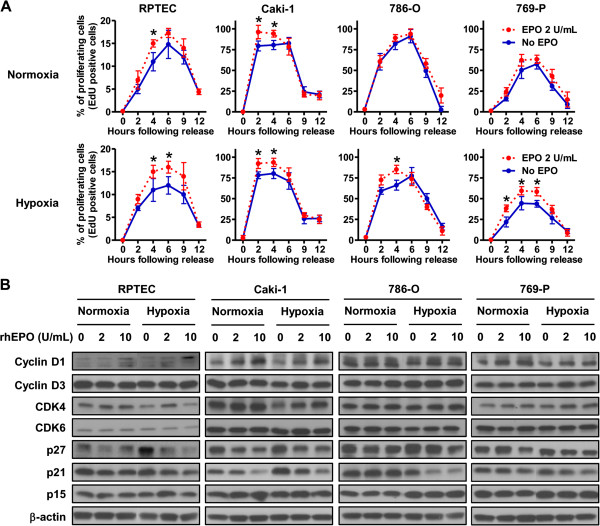
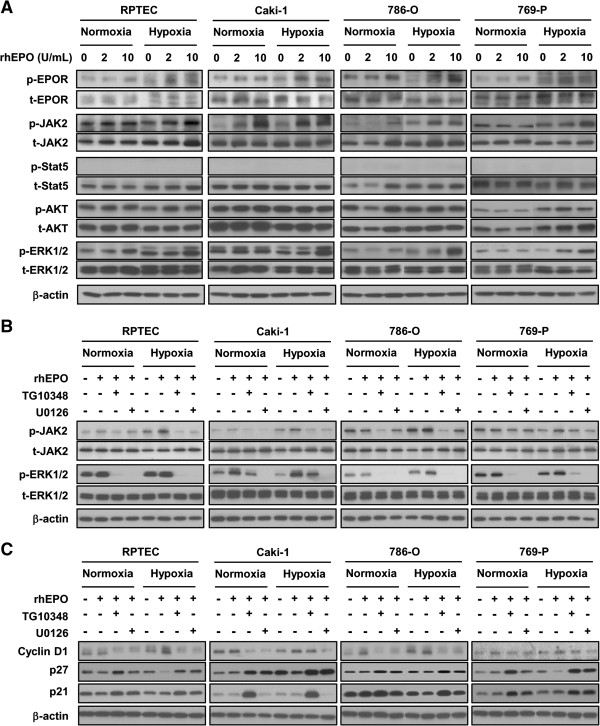
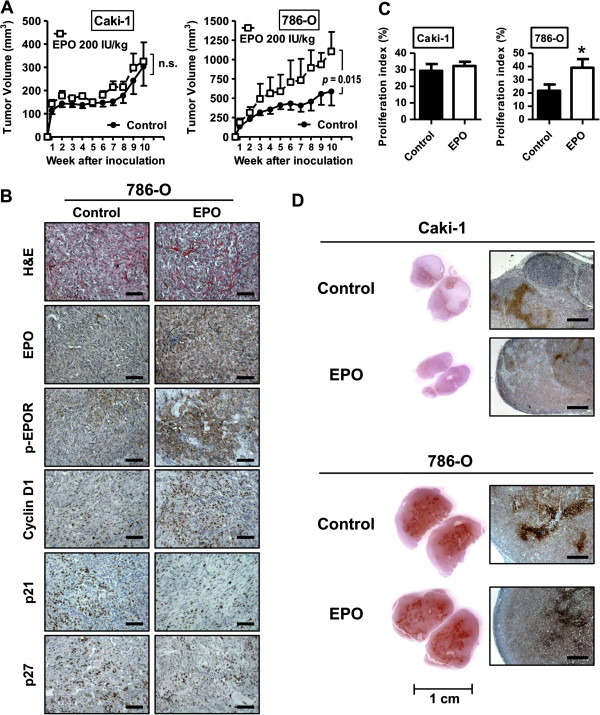
Results: EPO expression score was significantly elevated in lung cancer and lymphoma (compared to benign tissues), while EPOR expression score was significantly elevated in lymphoma, thyroid, uterine, lung and prostate cancers (compared to benign tissues). EPO and EPOR expression scores in RCC and benign renal tissue were not significantly different. Experimentally, we show that exposure of human renal cells to recombinant EPO (rhEPO) induces cellular proliferation, which we report for the first time, is further enhanced in a hypoxic state. Mechanistic investigations revealed that EPO stimulates the expression of cyclin D1 while inhibiting the expression of p21cip1 and p27kip1 through the phosphorylation of JAK2 and ERK1/2, leading to a more rapid progression through the cell cycle. We also demonstrate an increase in the growth of renal cell carcinoma xenograft tumors when systemic rhEPO is administered.
Conclusions: In summary, we elucidated a previously unidentified mechanism by which EPO administration regulates progression through the cell cycle, and show that EPO effects are significantly enhanced under hypoxic conditions.
Figures
References
-
- Krieg M, Haas R, Brauch H, Acker T, Flamme I, Plate KH. Up-regulation of hypoxia-inducible factors HIF-1alpha and HIF-2alpha under normoxic conditions in renal carcinoma cells by von Hippel-Lindau tumor suppressor gene loss of function. Oncogene. 2000;19:5435–5443. doi: 10.1038/sj.onc.1203938. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
