

The physics of functional magnetic resonance imaging (fMRI)
- PMID: 24006360
- PMCID: PMC4376284
- DOI: 10.1088/0034-4885/76/9/096601
The physics of functional magnetic resonance imaging (fMRI)
Abstract
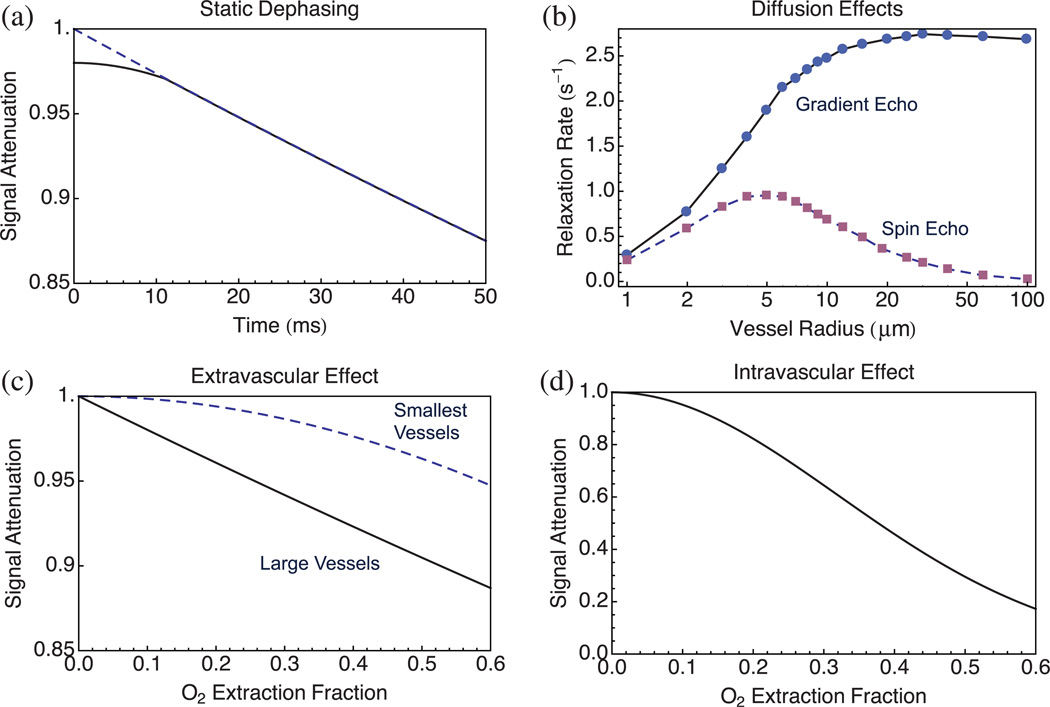
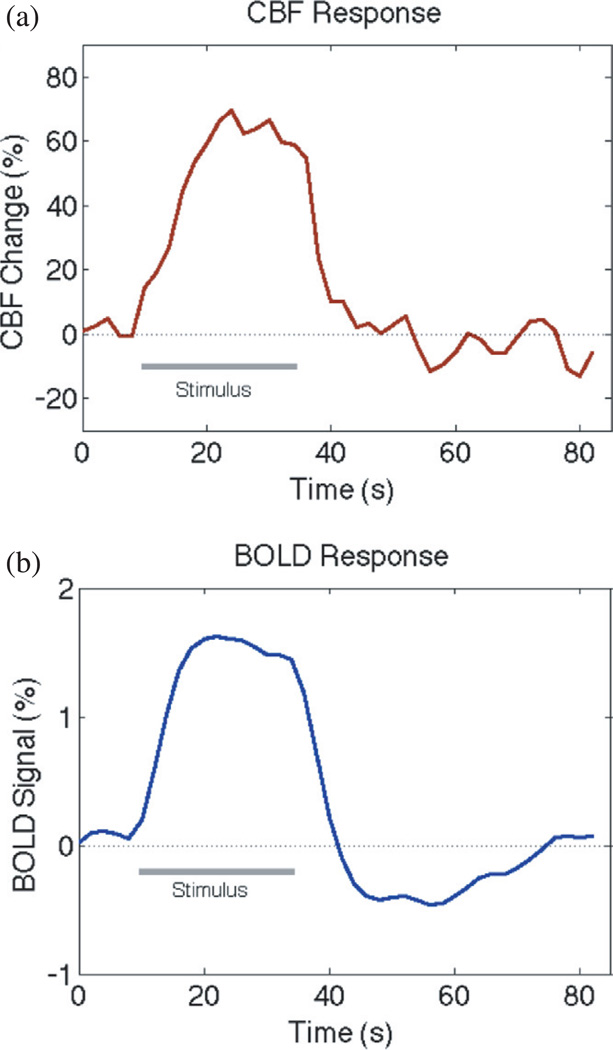
Functional magnetic resonance imaging (fMRI) is a methodology for detecting dynamic patterns of activity in the working human brain. Although the initial discoveries that led to fMRI are only about 20 years old, this new field has revolutionized the study of brain function. The ability to detect changes in brain activity has a biophysical basis in the magnetic properties of deoxyhemoglobin, and a physiological basis in the way blood flow increases more than oxygen metabolism when local neural activity increases. These effects translate to a subtle increase in the local magnetic resonance signal, the blood oxygenation level dependent (BOLD) effect, when neural activity increases. With current techniques, this pattern of activation can be measured with resolution approaching 1 mm(3) spatially and 1 s temporally. This review focuses on the physical basis of the BOLD effect, the imaging methods used to measure it, the possible origins of the physiological effects that produce a mismatch of blood flow and oxygen metabolism during neural activation, and the mathematical models that have been developed to understand the measured signals. An overarching theme is the growing field of quantitative fMRI, in which other MRI methods are combined with BOLD methods and analyzed within a theoretical modeling framework to derive quantitative estimates of oxygen metabolism and other physiological variables. That goal is the current challenge for fMRI: to move fMRI from a mapping tool to a quantitative probe of brain physiology.
Figures









References
-
- Bandettini PA. Twenty years of functional MRI: the science and the stories. Neuroimage. 2012;62:575–588. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical