

Rice CONSTITUTIVE TRIPLE-RESPONSE2 is involved in the ethylene-receptor signalling and regulation of various aspects of rice growth and development
- PMID: 24006427
- PMCID: PMC3830475
- DOI: 10.1093/jxb/ert272
Rice CONSTITUTIVE TRIPLE-RESPONSE2 is involved in the ethylene-receptor signalling and regulation of various aspects of rice growth and development
Abstract
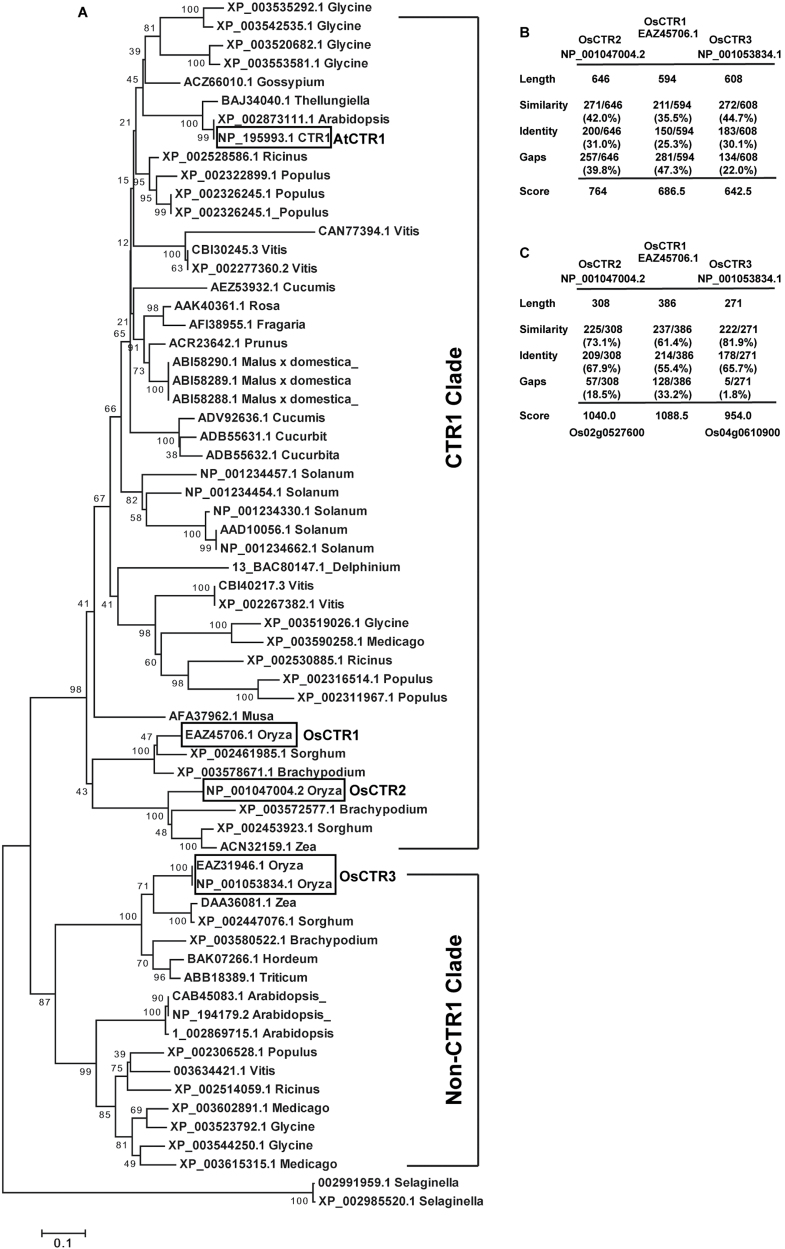
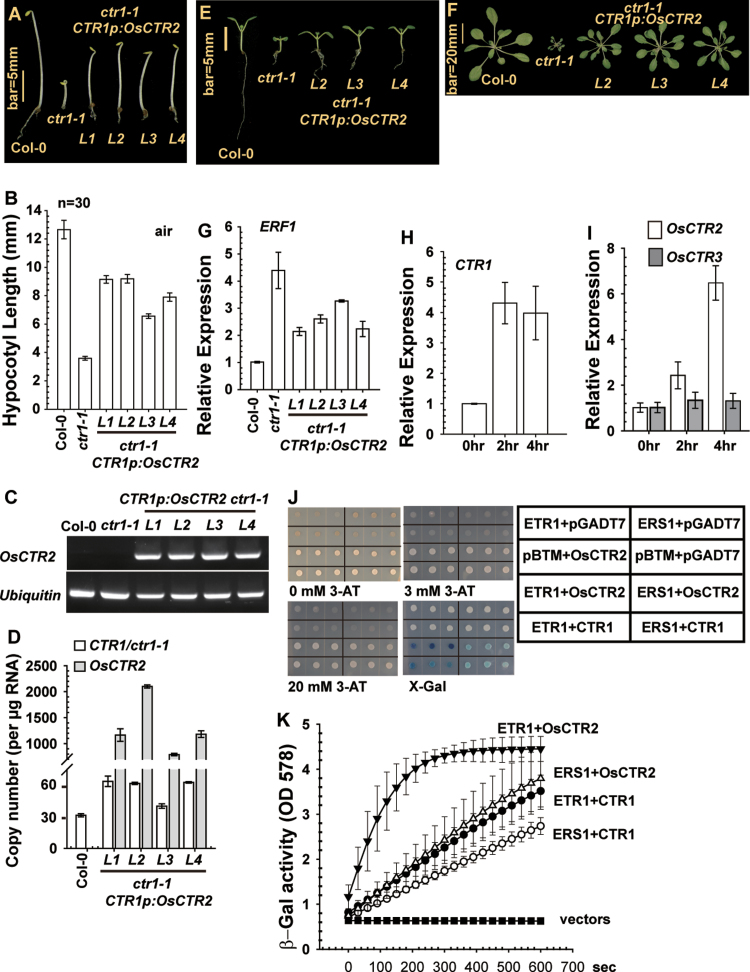
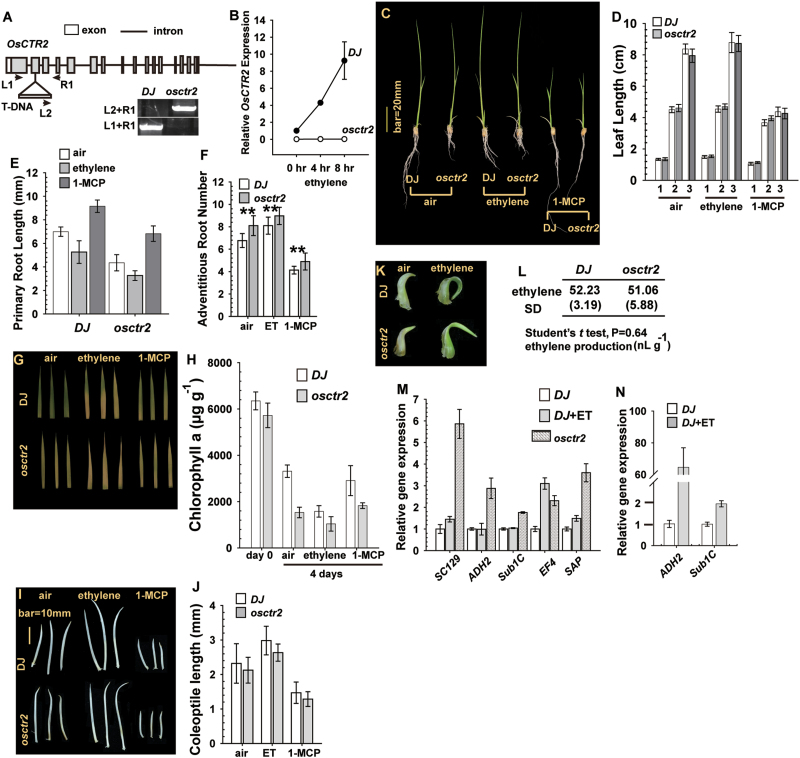
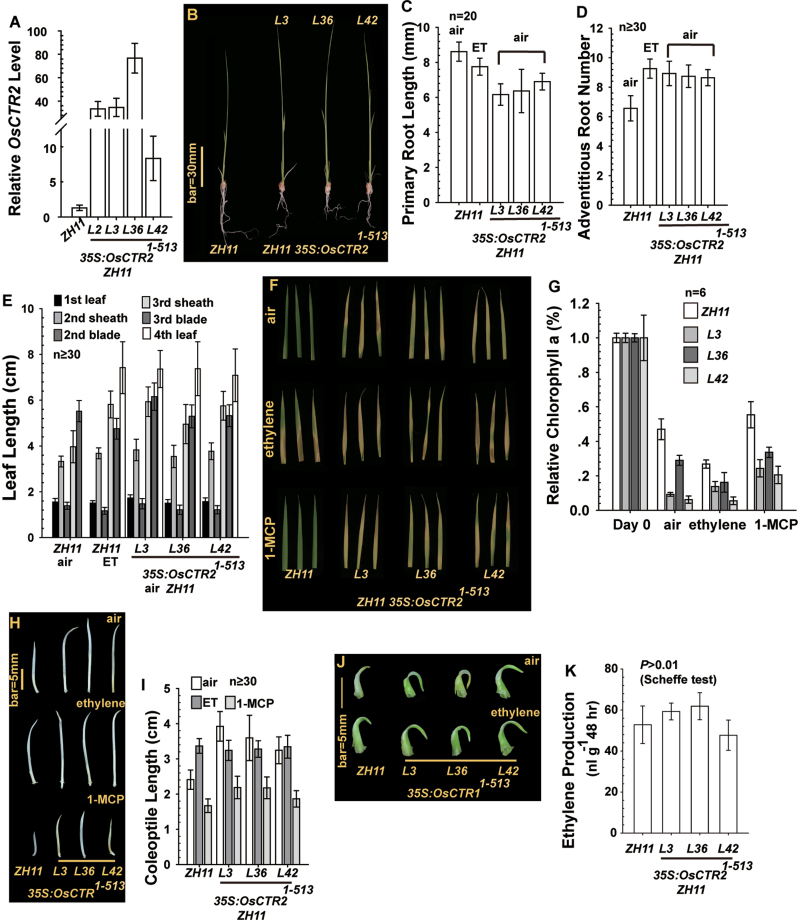
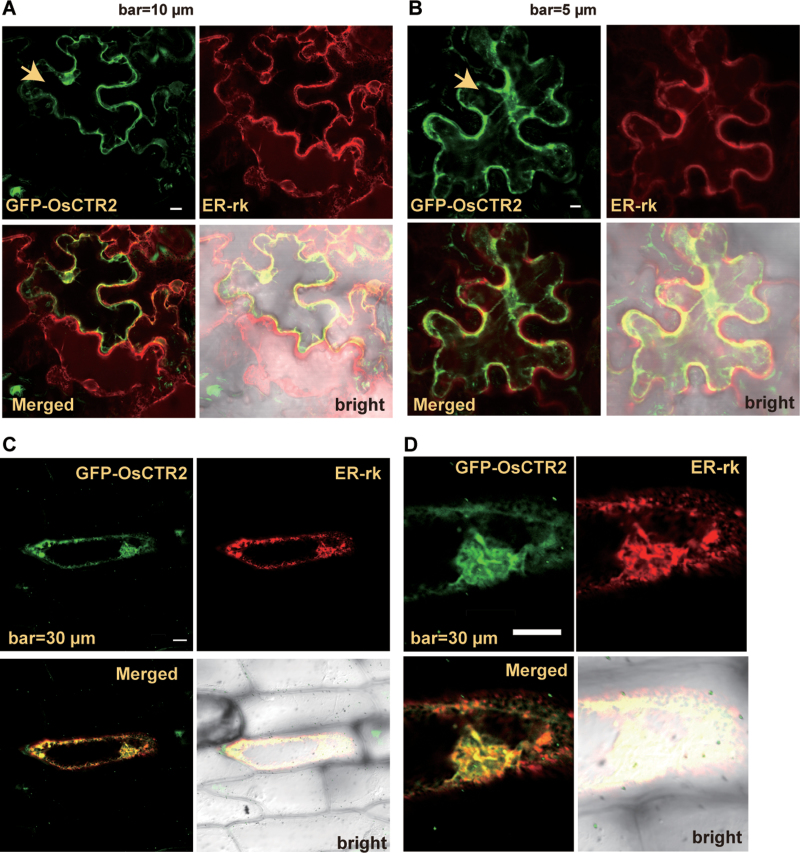
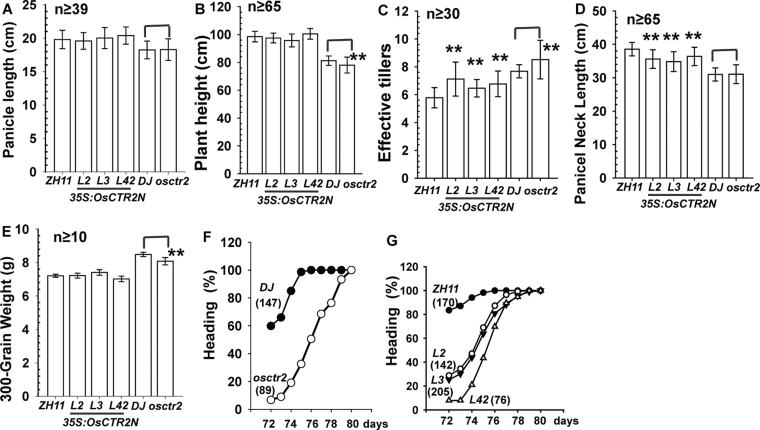
In Arabidopsis, the ethylene-receptor signal output occurs at the endoplasmic reticulum and is mediated by the Raf-like protein CONSTITUTIVE TRIPLE RESPONSE1 (CTR1) but is prevented by overexpression of the CTR1 N terminus. A phylogenic analysis suggested that rice OsCTR2 is closely related to CTR1, and ectopic expression of CTR1p:OsCTR2 complemented Arabidopsis ctr1-1. Arabidopsis ethylene receptors ETHYLENE RESPONSE1 and ETHYLENE RESPONSE SENSOR1 physically interacted with OsCTR2 on yeast two-hybrid assay, and green fluorescence protein-tagged OsCTR2 was localized at the endoplasmic reticulum. The osctr2 loss-of-function mutation and expression of the 35S:OsCTR2 (1-513) transgene that encodes the OsCTR2 N terminus (residues 1-513) revealed several and many aspects, respectively, of ethylene-induced growth alteration in rice. Because the osctr2 allele did not produce all aspects of ethylene-induced growth alteration, the ethylene-receptor signal output might be mediated in part by OsCTR2 and by other components in rice. Yield-related agronomic traits, including flowering time and effective tiller number, were altered in osctr2 and 35S:OsCTR2 (1-513) transgenic lines. Applying prolonged ethylene treatment to evaluate ethylene effects on rice without compromising rice growth is technically challenging. Our understanding of roles of ethylene in various aspects of growth and development in japonica rice varieties could be advanced with the use of the osctr2 and 35S:OsCTR2 (1-513) transgenic lines.
Figures
References
-
- Alexander L, Grierson D. 2002. Ethylene biosynthesis and action in tomato: a model for climacteric fruit ripening. Journal of Experimental Botany 53, 2039–2055 - PubMed
-
- Anderson JP, Badruzsaufari E, Schenk PM, Manners JM, Desmond OJ, Ehlert C, Maclean DJ, Ebert PR, Kazan K. 2004. Antagonistic interaction between abscisic acid and jasmonate-ethylene signaling pathways modulates defense gene expression and disease resistance in Arabidopsis. Plant Cell 16, 3460–3479 - PMC - PubMed
-
- Fukao T, Bailey-Serres J. 2008a. Ethylene—a key regulator of submergence responses in rice. Plant Science 175, 43–51
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
