

Differential 12-O-Tetradecanoylphorbol-13-acetate-induced activation of rat mammary carcinoma susceptibility Fbxo10 variant promoters via a PKC-AP1 pathway
- PMID: 24008983
- PMCID: PMC9733134
- DOI: 10.1002/mc.22081
Differential 12-O-Tetradecanoylphorbol-13-acetate-induced activation of rat mammary carcinoma susceptibility Fbxo10 variant promoters via a PKC-AP1 pathway
Abstract
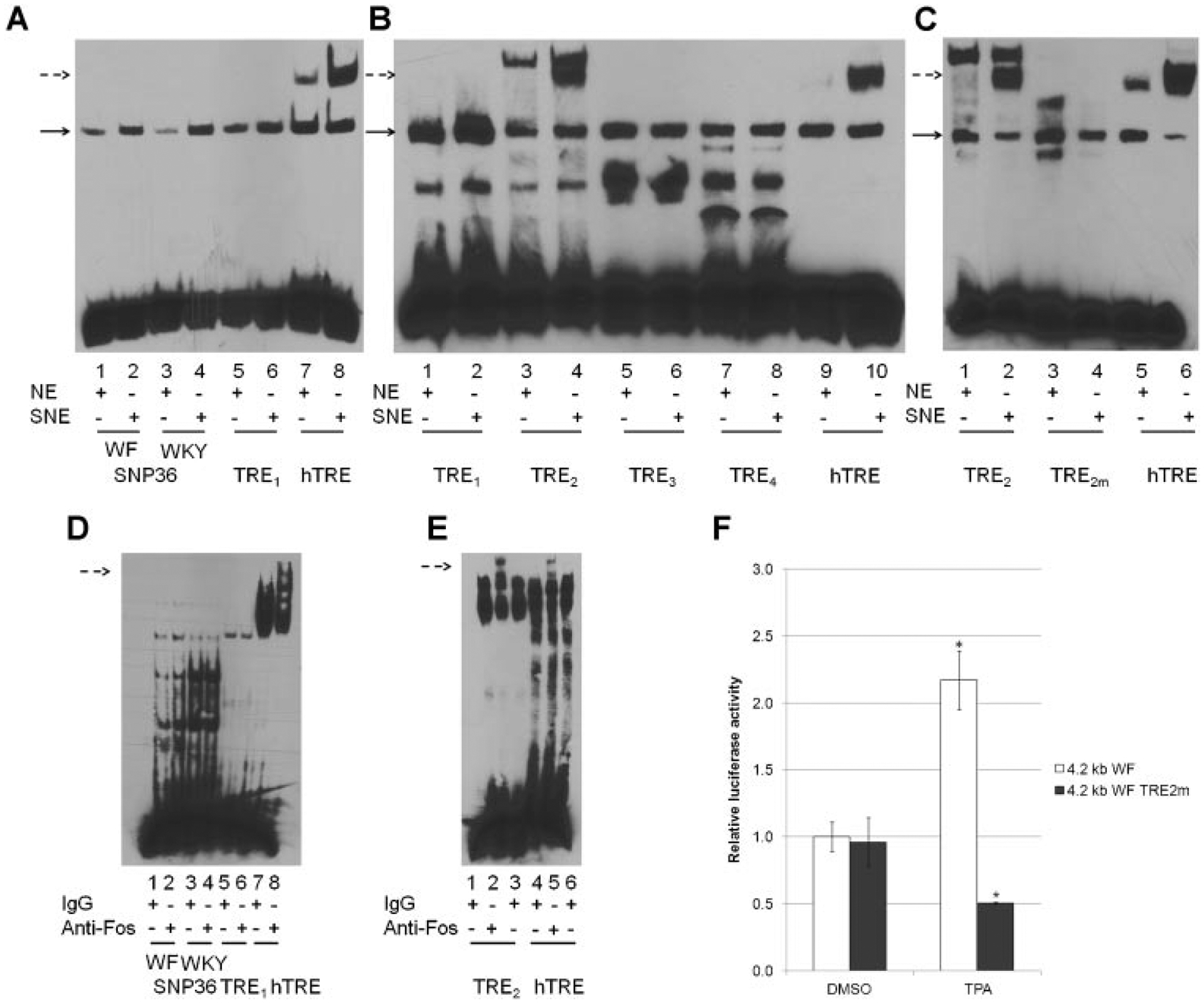
Rat mammary carcinoma susceptibility 5a1 (Mcs5a1), which is concordant to human MCS5A1 breast cancer risk locus, mediates susceptibility by a non-mammary cell-autonomous mechanism associated with T cell differential expression of F-box protein 10 (Fbxo10). Human FBXO10, an evolutionarily conserved ubiquitin ligase gene, was shown to have a potential role in regulating cell death by controlling the degradation of Bcl-2, a key protein involved in apoptosis. Breast cancer susceptibility is controlled by interactions between environmental and genetic factors; therefore, we sought to determine if breast cancer risk-associated environmental chemicals interact with Mcs5a1 variants using luciferase reporter constructs containing 4.2 kb Fbxo10 promoters based on alleles of mammary cancer susceptible Wistar Furth (WF) and resistant Wistar Kyoto (WKY) rat strains. 12-O-Tetradecanoylphorbol-13-acetate (TPA) induced activation of a 4.2 kb WF Fbxo10 promoter region, but lower levels of activation of the homologous WKY Fbxo10 promoter region. Using general and specific protein kinase inhibitors, we identified a protein kinase C (PKC) pathway that mediated TPA activation. We narrowed the possible PKCs to a member of the atypical PKC isoforms, namely PKCµ. We also determined that activator protein 1 (AP1) family member c-Fos mediated TPA activation of the 4.2 kb WF Fbxo10 promoter. TPA was shown to induce endogenous FBXO10 mRNA and FBXO10 protein in Jurkat cells, a human T cell line, with a maximal level of expression from 1.5 to 2.5 h after exposure. These results indicate that FBXO10/Fbxo10 expression is regulated by a PKC-dependent pathway acting through c-Fos, which binds AP1-specific DNA elements in Mcs5a1.
Keywords: F-box protein 10; activator protein 1; breast cancer susceptibility; mammary carcinoma susceptibility 5a1; protein kinase C.
© 2013 Wiley Periodicals, Inc.
Figures






References
-
- Siegel R, Naishadham D, Jemal A. Cancer statistics. CA Cancer J Clin 2012;62:10–29. - PubMed
-
- Bond GL. Hu W. Bond EE, et al. A single nucleotide polymorphism in the MDM2 promoter attenuates the p53 tumor suppressor pathway and accelerates tumor formation in humans. Cell 2004;119:591–602. - PubMed
-
- Wu AH, Wan P, Hankin J, et al. Adolescent and adult soy intake and risk of breast cancer in Asian-Americans. Carcinogenesis 2002;23:1491–1496. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
