

Recurrent modification of a conserved cis-regulatory element underlies fruit fly pigmentation diversity
- PMID: 24009528
- PMCID: PMC3757066
- DOI: 10.1371/journal.pgen.1003740
Recurrent modification of a conserved cis-regulatory element underlies fruit fly pigmentation diversity
Abstract
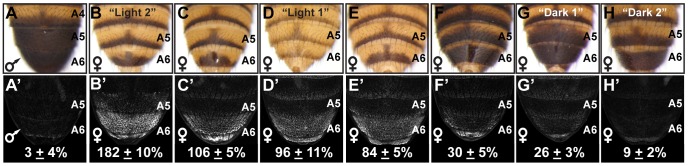
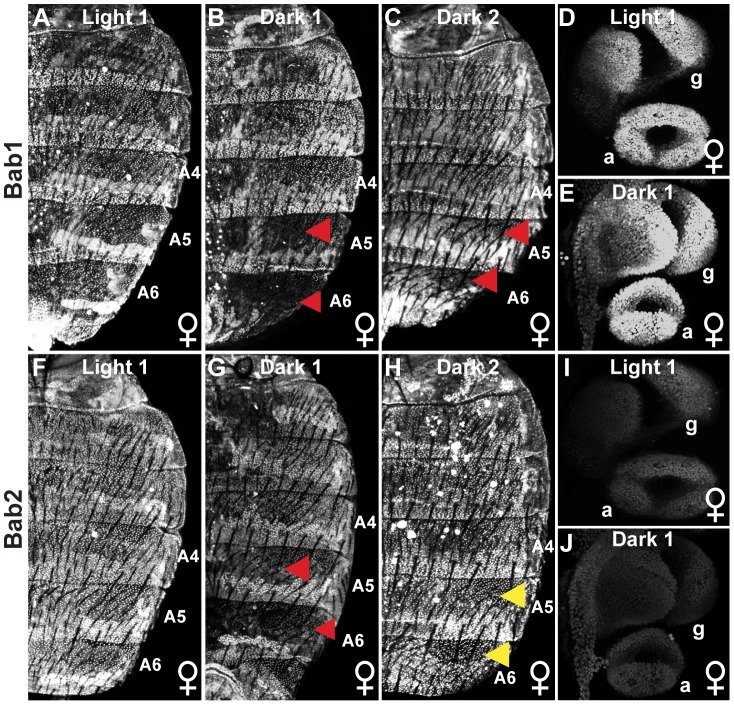
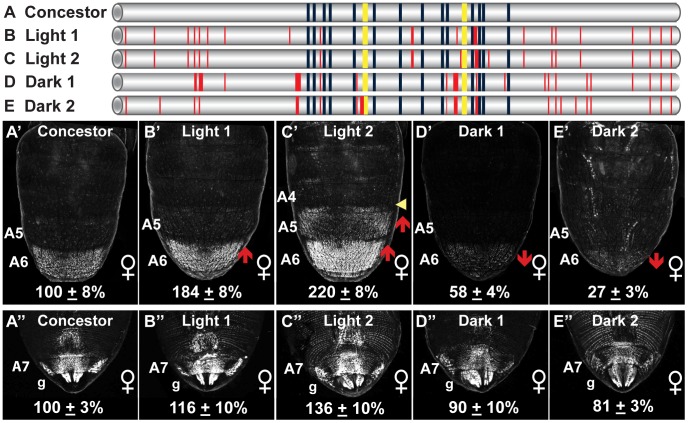
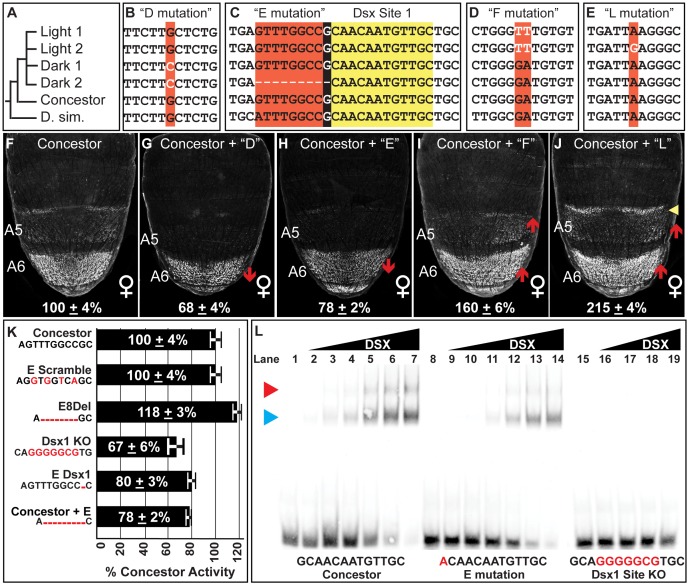
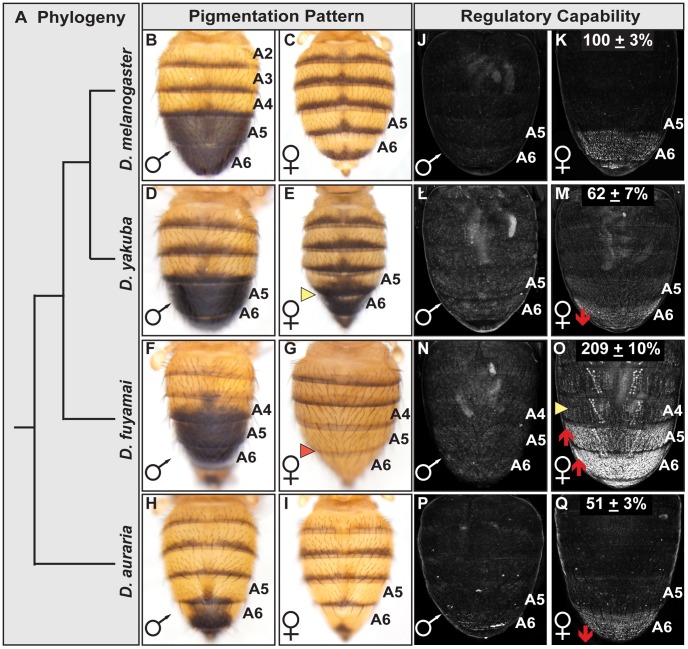
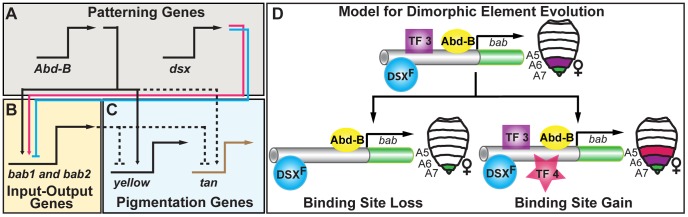
The development of morphological traits occurs through the collective action of networks of genes connected at the level of gene expression. As any node in a network may be a target of evolutionary change, the recurrent targeting of the same node would indicate that the path of evolution is biased for the relevant trait and network. Although examples of parallel evolution have implicated recurrent modification of the same gene and cis-regulatory element (CRE), little is known about the mutational and molecular paths of parallel CRE evolution. In Drosophila melanogaster fruit flies, the Bric-à-brac (Bab) transcription factors control the development of a suite of sexually dimorphic traits on the posterior abdomen. Female-specific Bab expression is regulated by the dimorphic element, a CRE that possesses direct inputs from body plan (ABD-B) and sex-determination (DSX) transcription factors. Here, we find that the recurrent evolutionary modification of this CRE underlies both intraspecific and interspecific variation in female pigmentation in the melanogaster species group. By reconstructing the sequence and regulatory activity of the ancestral Drosophila melanogaster dimorphic element, we demonstrate that a handful of mutations were sufficient to create independent CRE alleles with differing activities. Moreover, intraspecific and interspecific dimorphic element evolution proceeded with little to no alterations to the known body plan and sex-determination regulatory linkages. Collectively, our findings represent an example where the paths of evolution appear biased to a specific CRE, and drastic changes in function were accompanied by deep conservation of key regulatory linkages.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures

References
-
- Conway Morris S (2003) Life's Solution. Inevitable Humans in a Lonely Universe. 1st ed. Cambridge: Cambridge University Press.
-
- Gompel N, Prud'homme B (2009) The causes of repeated genetic evolution. Developmental biology 332: 36–47 Available:http://www.ncbi.nlm.nih.gov/pubmed/19433086. Accessed 12 March 2013. - PubMed
-
- Shapiro MD, Bell MA, Kingsley DM (2006) Parallel genetic origins of pelvic reduction in vertebrates. Proceedings of the National Academy of Sciences of the United States of America 103: 13753–13758 http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=1564237&tool=p.... - PMC - PubMed
-
- Protas ME, Hersey C, Kochanek D, Zhou Y, Wilkens H, et al. (2006) Genetic analysis of cavefish reveals molecular convergence in the evolution of albinism. Nature genetics 38: 107–111 Available:http://www.ncbi.nlm.nih.gov/pubmed/16341223. Accessed 1 March 2013. - PubMed
-
- Sucena E, Delon I, Jones I, Payre F, Stern DL (2003) Regulatory evolution of shavenbaby/ovo underlies multiple cases of morphological parallelism. Nature 424: 935–938 Available:http://www.ncbi.nlm.nih.gov/pubmed/12931187. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
