

Are vesicular neurotransmitter transporters potential treatment targets for temporal lobe epilepsy?
- PMID: 24009559
- PMCID: PMC3757300
- DOI: 10.3389/fncel.2013.00139
Are vesicular neurotransmitter transporters potential treatment targets for temporal lobe epilepsy?
Abstract
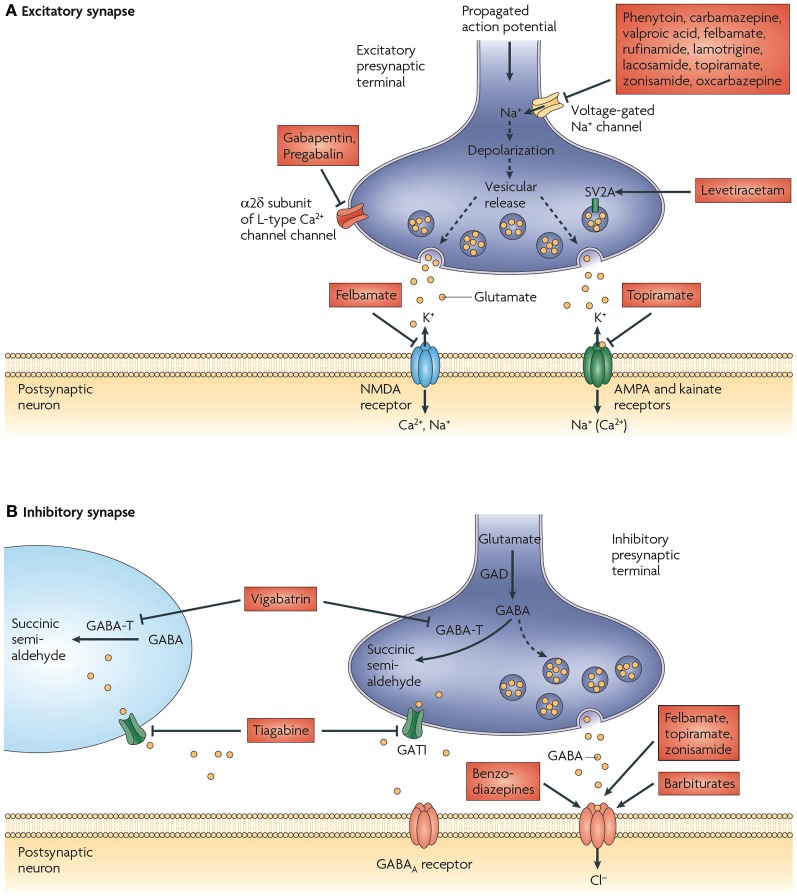
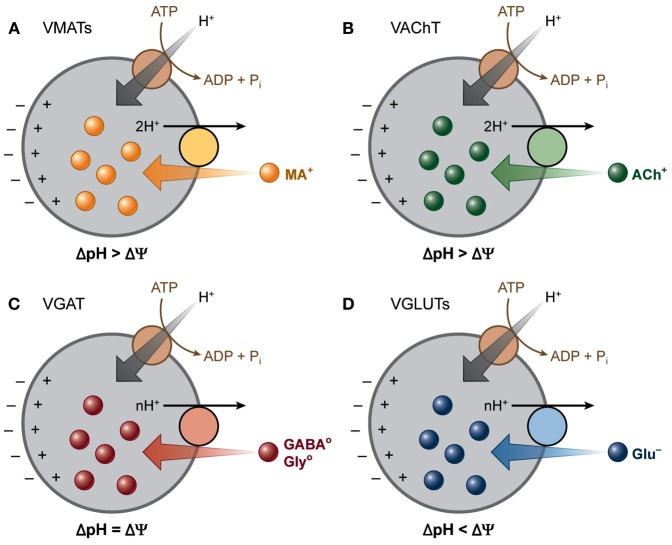
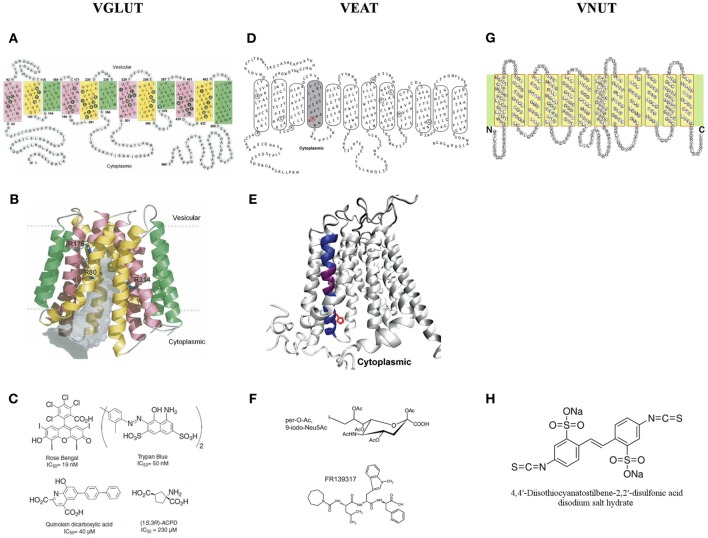
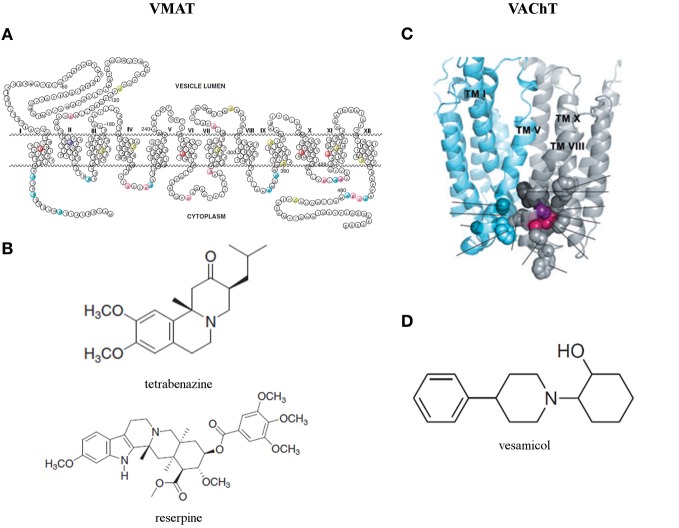
The vesicular neurotransmitter transporters (VNTs) are small proteins responsible for packing synaptic vesicles with neurotransmitters thereby determining the amount of neurotransmitter released per vesicle through fusion in both neurons and glial cells. Each transporter subtype was classically seen as a specific neuronal marker of the respective nerve cells containing that particular neurotransmitter or structurally related neurotransmitters. More recently, however, it has become apparent that common neurotransmitters can also act as co-transmitters, adding complexity to neurotransmitter release and suggesting intriguing roles for VNTs therein. We will first describe the current knowledge on vesicular glutamate transporters (VGLUT1/2/3), the vesicular excitatory amino acid transporter (VEAT), the vesicular nucleotide transporter (VNUT), vesicular monoamine transporters (VMAT1/2), the vesicular acetylcholine transporter (VAChT) and the vesicular γ-aminobutyric acid (GABA) transporter (VGAT) in the brain. We will focus on evidence regarding transgenic mice with disruptions in VNTs in different models of seizures and epilepsy. We will also describe the known alterations and reorganizations in the expression levels of these VNTs in rodent models for temporal lobe epilepsy (TLE) and in human tissue resected for epilepsy surgery. Finally, we will discuss perspectives on opportunities and challenges for VNTs as targets for possible future epilepsy therapies.
Keywords: SLC17; SLC18; SLC32; antiepileptic drugs; epileptogenesis; temporal lobe epilepsy; vesicular neurotransmitter transporters.
Figures
References
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources