

An X-linked cobalamin disorder caused by mutations in transcriptional coregulator HCFC1
- PMID: 24011988
- PMCID: PMC3769968
- DOI: 10.1016/j.ajhg.2013.07.022
An X-linked cobalamin disorder caused by mutations in transcriptional coregulator HCFC1
Abstract
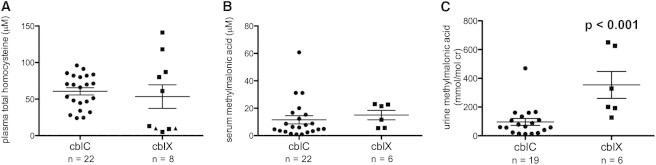
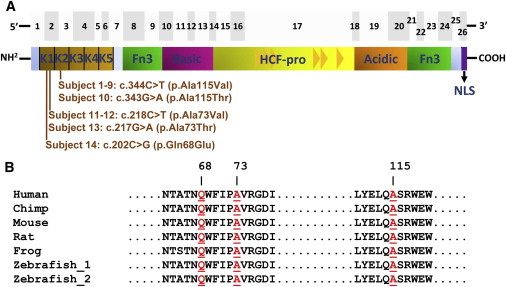
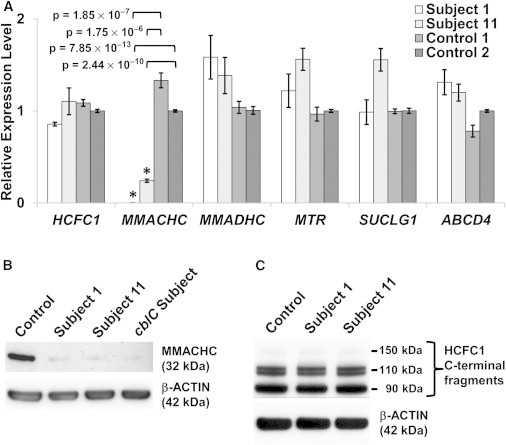
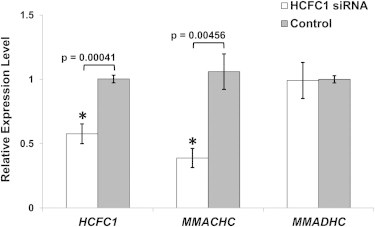
Derivatives of vitamin B12 (cobalamin) are essential cofactors for enzymes required in intermediary metabolism. Defects in cobalamin metabolism lead to disorders characterized by the accumulation of methylmalonic acid and/or homocysteine in blood and urine. The most common inborn error of cobalamin metabolism, combined methylmalonic acidemia and hyperhomocysteinemia, cblC type, is caused by mutations in MMACHC. However, several individuals with presumed cblC based on cellular and biochemical analysis do not have mutations in MMACHC. We used exome sequencing to identify the genetic basis of an X-linked form of combined methylmalonic acidemia and hyperhomocysteinemia, designated cblX. A missense mutation in a global transcriptional coregulator, HCFC1, was identified in the index case. Additional male subjects were ascertained through two international diagnostic laboratories, and 13/17 had one of five distinct missense mutations affecting three highly conserved amino acids within the HCFC1 kelch domain. A common phenotype of severe neurological symptoms including intractable epilepsy and profound neurocognitive impairment, along with variable biochemical manifestations, was observed in all affected subjects compared to individuals with early-onset cblC. The severe reduction in MMACHC mRNA and protein within subject fibroblast lines suggested a role for HCFC1 in transcriptional regulation of MMACHC, which was further supported by the identification of consensus HCFC1 binding sites in MMACHC. Furthermore, siRNA-mediated knockdown of HCFC1 expression resulted in the coordinate downregulation of MMACHC mRNA. This X-linked disorder demonstrates a distinct disease mechanism by which transcriptional dysregulation leads to an inborn error of metabolism with a complex clinical phenotype.
Copyright © 2013 The American Society of Human Genetics. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Watkins D., Rosenblatt D.S. Inborn errors of cobalamin absorption and metabolism. Am. J. Med. Genet. C. Semin. Med. Genet. 2011;157:33–44. - PubMed
-
- Lerner-Ellis J.P., Anastasio N., Liu J., Coelho D., Suormala T., Stucki M., Loewy A.D., Gurd S., Grundberg E., Morel C.F. Spectrum of mutations in MMACHC, allelic expression, and evidence for genotype-phenotype correlations. Hum. Mutat. 2009;30:1072–1081. - PubMed
-
- Martinelli D., Deodato F., Dionisi-Vici C. Cobalamin C defect: natural history, pathophysiology, and treatment. J. Inherit. Metab. Dis. 2011;34:127–135. - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
