

A cancer-associated BRCA2 mutation reveals masked nuclear export signals controlling localization
- PMID: 24013206
- PMCID: PMC3796201
- DOI: 10.1038/nsmb.2666
A cancer-associated BRCA2 mutation reveals masked nuclear export signals controlling localization
Abstract
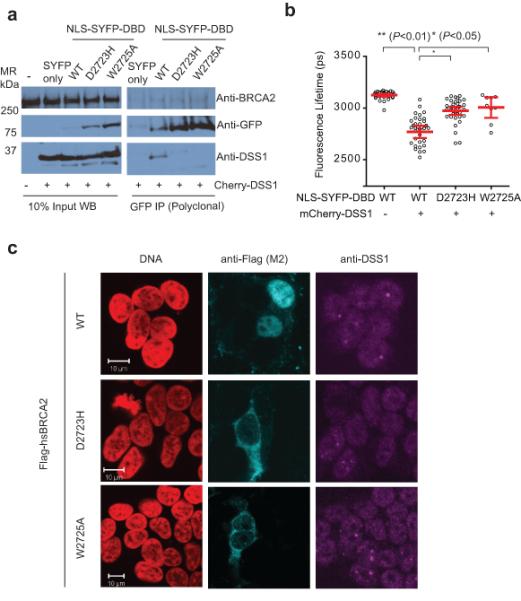
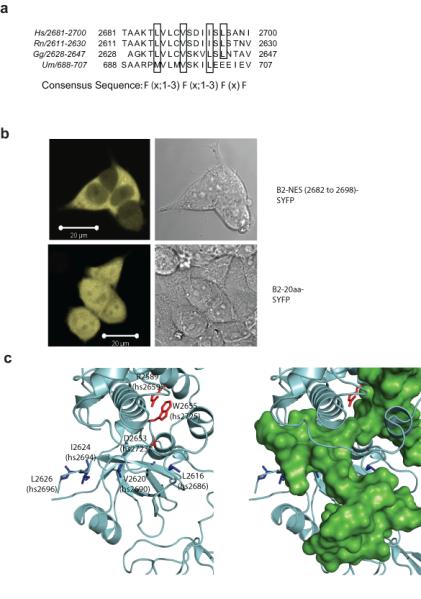
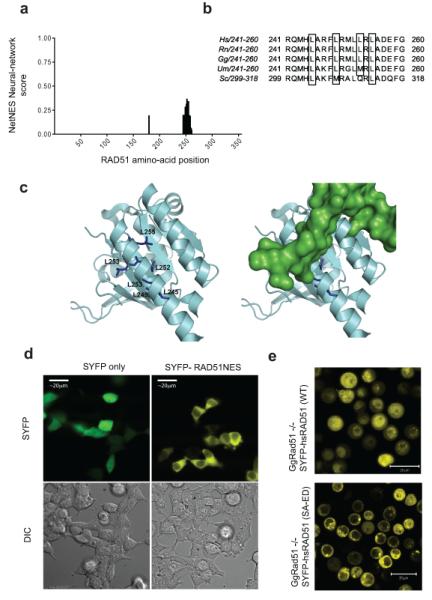
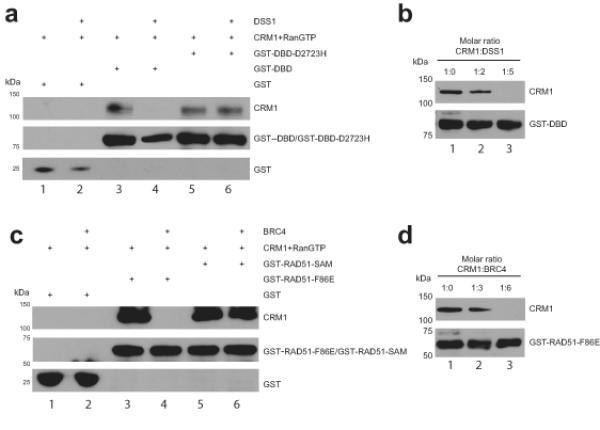
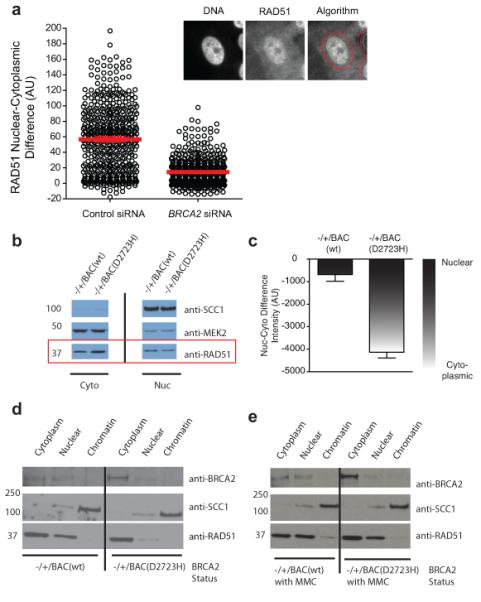
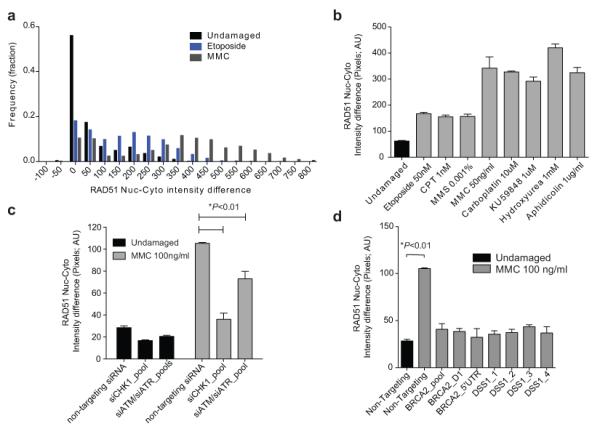
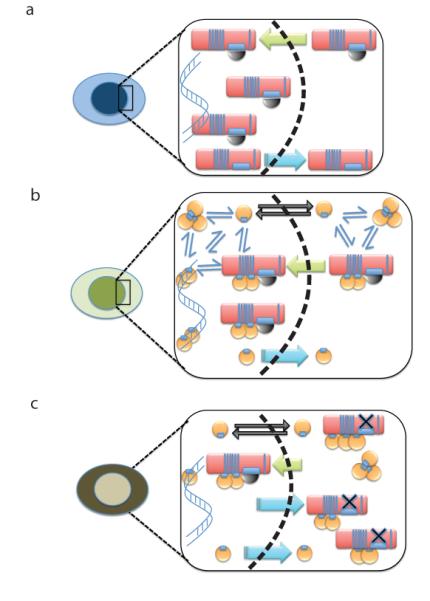
Germline missense mutations affecting a single BRCA2 allele predispose humans to cancer. Here we identify a protein-targeting mechanism that is disrupted by the cancer-associated mutation, BRCA2(D2723H), and that controls the nuclear localization of BRCA2 and its cargo, the recombination enzyme RAD51. A nuclear export signal (NES) in BRCA2 is masked by its interaction with a partner protein, DSS1, such that point mutations impairing BRCA2-DSS1 binding render BRCA2 cytoplasmic. In turn, cytoplasmic mislocalization of mutant BRCA2 inhibits the nuclear retention of RAD51 by exposing a similar NES in RAD51 that is usually obscured by the BRCA2-RAD51 interaction. Thus, a series of NES-masking interactions localizes BRCA2 and RAD51 in the nucleus. Notably, BRCA2(D2723H) decreases RAD51 nuclear retention even when wild-type BRCA2 is also present. Our findings suggest a mechanism for the regulation of the nucleocytoplasmic distribution of BRCA2 and RAD51 and its impairment by a heterozygous disease-associated mutation.
Figures
Similar articles
-
Promotion of BRCA2-Dependent Homologous Recombination by DSS1 via RPA Targeting and DNA Mimicry.Mol Cell. 2015 Jul 16;59(2):176-87. doi: 10.1016/j.molcel.2015.05.032. Epub 2015 Jul 2. Mol Cell. 2015. PMID: 26145171 Free PMC article.
-
DSS1 restrains BRCA2's engagement with dsDNA for homologous recombination, replication fork protection, and R-loop homeostasis.Nat Commun. 2024 Aug 17;15(1):7081. doi: 10.1038/s41467-024-51557-6. Nat Commun. 2024. PMID: 39152168 Free PMC article.
-
The carboxyl terminus of Brca2 links the disassembly of Rad51 complexes to mitotic entry.Curr Biol. 2009 Jul 14;19(13):1075-85. doi: 10.1016/j.cub.2009.05.057. Epub 2009 Jun 18. Curr Biol. 2009. PMID: 19540122 Free PMC article.
-
BRCA2: one small step for DNA repair, one giant protein purified.Yale J Biol Med. 2013 Dec 13;86(4):479-89. Yale J Biol Med. 2013. PMID: 24348212 Free PMC article. Review.
-
DSS1/Sem1, a Multifunctional and Intrinsically Disordered Protein.Trends Biochem Sci. 2016 May;41(5):446-459. doi: 10.1016/j.tibs.2016.02.004. Epub 2016 Mar 1. Trends Biochem Sci. 2016. PMID: 26944332 Review.
Cited by
-
Novel insights into RAD51 activity and regulation during homologous recombination and DNA replication.Biochem Cell Biol. 2016 Oct;94(5):407-418. doi: 10.1139/bcb-2016-0012. Epub 2016 Mar 31. Biochem Cell Biol. 2016. PMID: 27224545 Free PMC article. Review.
-
Promotion of BRCA2-Dependent Homologous Recombination by DSS1 via RPA Targeting and DNA Mimicry.Mol Cell. 2015 Jul 16;59(2):176-87. doi: 10.1016/j.molcel.2015.05.032. Epub 2015 Jul 2. Mol Cell. 2015. PMID: 26145171 Free PMC article.
-
Imprecise Medicine: BRCA2 Variants of Uncertain Significance (VUS), the Challenges and Benefits to Integrate a Functional Assay Workflow with Clinical Decision Rules.Genes (Basel). 2021 May 20;12(5):780. doi: 10.3390/genes12050780. Genes (Basel). 2021. PMID: 34065235 Free PMC article. Review.
-
The Pathogenic R3052W BRCA2 Variant Disrupts Homology-Directed Repair by Failing to Localize to the Nucleus.Front Genet. 2022 May 30;13:884210. doi: 10.3389/fgene.2022.884210. eCollection 2022. Front Genet. 2022. PMID: 35711920 Free PMC article.
-
GFP-Fragment Reassembly Screens for the Functional Characterization of Variants of Uncertain Significance in Protein Interaction Domains of the BRCA1 and BRCA2 Genes.Cancers (Basel). 2019 Jan 28;11(2):151. doi: 10.3390/cancers11020151. Cancers (Basel). 2019. PMID: 30696104 Free PMC article.
References
-
- The Breast Cancer Linkage Consortium Cancer risks in BRCA2 mutation carriers. The Breast Cancer Linkage Consortium. J Natl Cancer Inst. 1999;91:1310–6. - PubMed
-
- Collins N, et al. Consistent loss of the wild type allele in breast cancers from a family linked to the BRCA2 gene on chromosome 13q12-13. Oncogene. 1995;10:1673–5. - PubMed
-
- King TA, et al. Heterogenic loss of the wild-type BRCA allele in human breast tumorigenesis. Ann Surg Oncol. 2007;14:2510–8. - PubMed
-
- Skoulidis F, et al. Germline Brca2 heterozygosity promotes Kras(G12D) -driven carcinogenesis in a murine model of familial pancreatic cancer. Cancer Cell. 2010;18:499–509. - PubMed
References for Methods
-
- Yu DS, et al. Dynamic control of Rad51 recombinase by self-association and interaction with BRCA2. Mol Cell. 2003;12:1029–41. - PubMed
-
- Guttler T, et al. NES consensus redefined by structures of PKI-type and Rev-type nuclear export signals bound to CRM1. Nat Struct Mol Biol. 2010;17:1367–1376. - PubMed
-
- Frey S, Richter RP, Gorlich D. FG-rich repeats of nuclear pore proteins form a three-dimensional meshwork with hydrogel-like properties. Science. 2006;314:815–7. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
