

RecG and UvsW catalyse robust DNA rewinding critical for stalled DNA replication fork rescue
- PMID: 24013402
- PMCID: PMC3778716
- DOI: 10.1038/ncomms3368
RecG and UvsW catalyse robust DNA rewinding critical for stalled DNA replication fork rescue
Erratum in
- Nat Commun. 2014;5:4210. Bianco, Piero [corrected to Bianco, Piero R]
Abstract
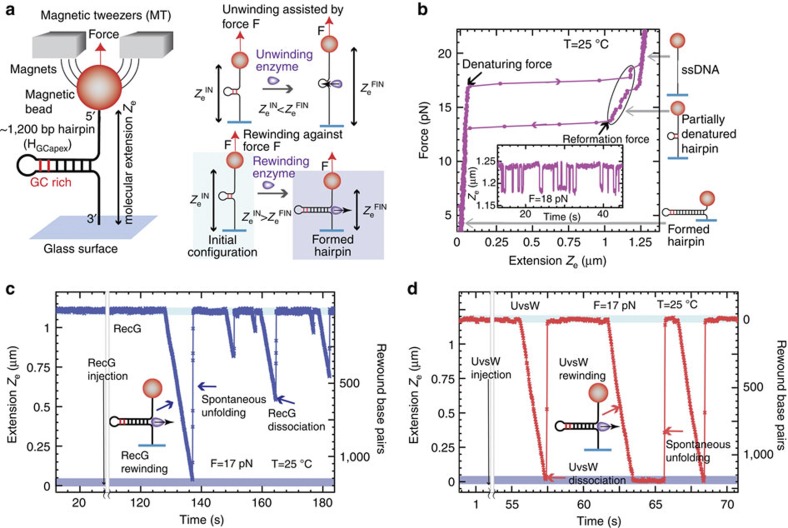
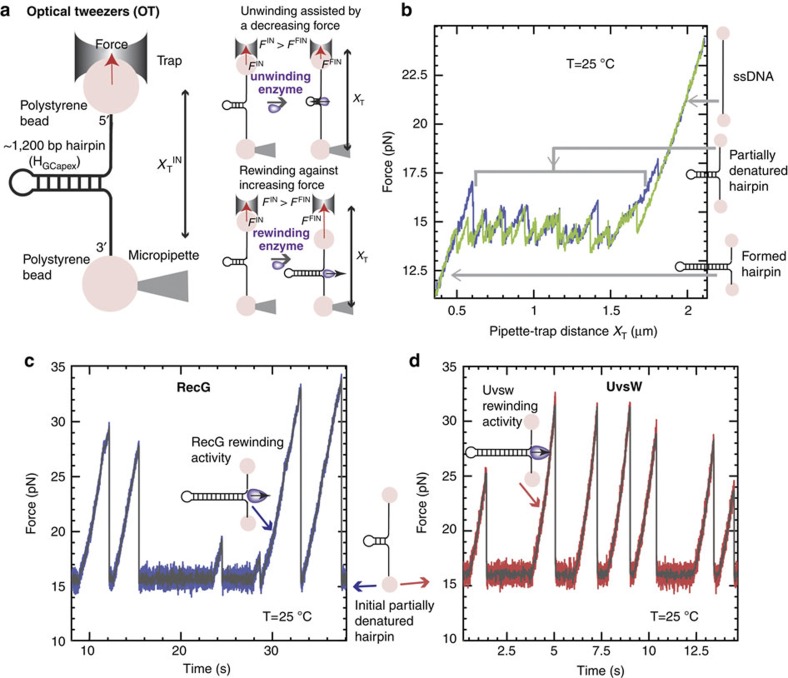
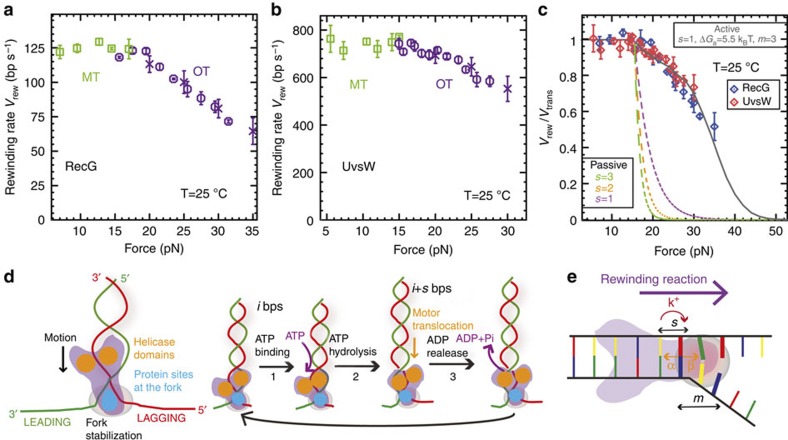
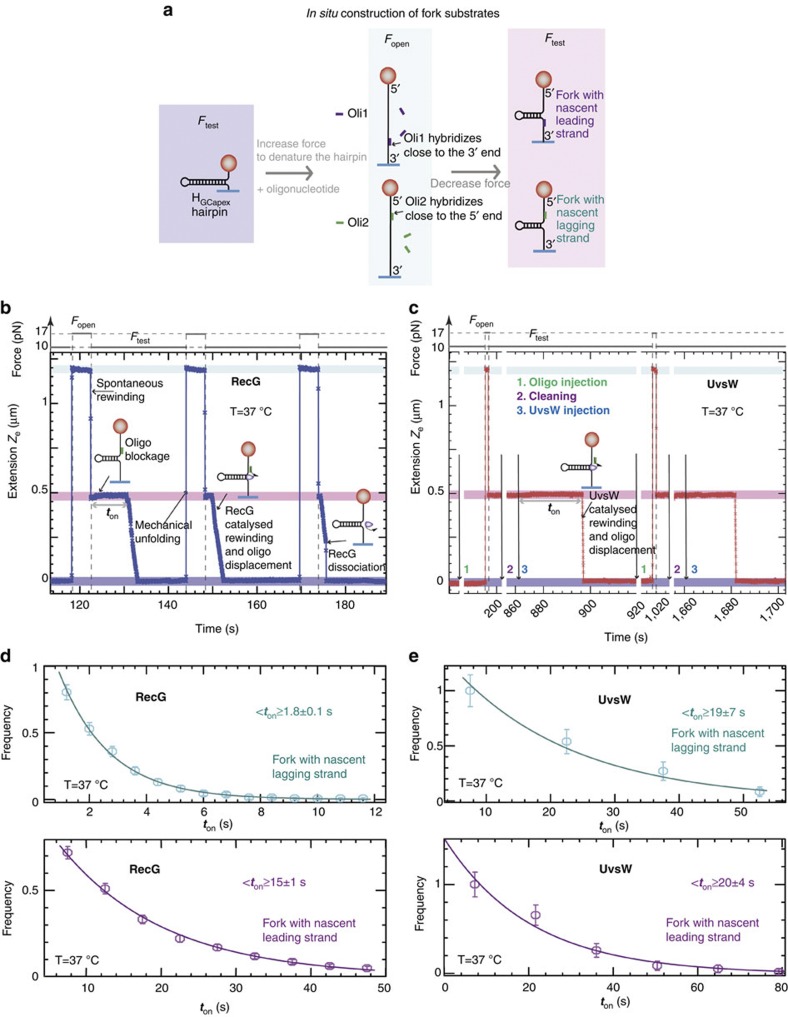
Helicases that both unwind and rewind DNA have central roles in DNA repair and genetic recombination. In contrast to unwinding, DNA rewinding by helicases has proved difficult to characterize biochemically because of its thermodynamically downhill nature. Here we use single-molecule assays to mechanically destabilize a DNA molecule and follow, in real time, unwinding and rewinding by two DNA repair helicases, bacteriophage T4 UvsW and Escherichia coli RecG. We find that both enzymes are robust rewinding enzymes, which can work against opposing forces as large as 35 pN, revealing their active character. The generation of work during the rewinding reaction allows them to couple rewinding to DNA unwinding and/or protein displacement reactions central to the rescue of stalled DNA replication forks. The overall results support a general mechanism for monomeric rewinding enzymes.
Figures
Similar articles
-
The Biochemical Mechanism of Fork Regression in Prokaryotes and Eukaryotes-A Single Molecule Comparison.Int J Mol Sci. 2022 Aug 3;23(15):8613. doi: 10.3390/ijms23158613. Int J Mol Sci. 2022. PMID: 35955746 Free PMC article. Review.
-
Stalled replication fork rescue requires a novel DNA helicase.Methods. 2016 Oct 1;108:40-7. doi: 10.1016/j.ymeth.2016.06.002. Epub 2016 Jun 6. Methods. 2016. PMID: 27282357 Free PMC article. Review.
-
Characterization of the ATPase activity of RecG and RuvAB proteins on model fork structures reveals insight into stalled DNA replication fork repair.J Biol Chem. 2013 Sep 13;288(37):26397-409. doi: 10.1074/jbc.M113.500223. Epub 2013 Jul 27. J Biol Chem. 2013. PMID: 23893472 Free PMC article.
-
Bacteriophage T4 UvsW protein is a helicase involved in recombination, repair and the regulation of DNA replication origins.EMBO J. 1997 Jul 1;16(13):4142-51. doi: 10.1093/emboj/16.13.4142. EMBO J. 1997. PMID: 9233823 Free PMC article.
-
Direct observation of stalled fork restart via fork regression in the T4 replication system.Science. 2012 Nov 30;338(6111):1217-20. doi: 10.1126/science.1225437. Science. 2012. PMID: 23197534 Free PMC article.
Cited by
-
Remodeling of RecG Helicase at the DNA Replication Fork by SSB Protein.Sci Rep. 2015 Apr 29;5:9625. doi: 10.1038/srep09625. Sci Rep. 2015. PMID: 25923319 Free PMC article.
-
Cellular location and activity of Escherichia coli RecG proteins shed light on the function of its structurally unresolved C-terminus.Nucleic Acids Res. 2014 May;42(9):5702-14. doi: 10.1093/nar/gku228. Epub 2014 Apr 1. Nucleic Acids Res. 2014. PMID: 24692661 Free PMC article.
-
Interaction of branch migration translocases with the Holliday junction-resolving enzyme and their implications in Holliday junction resolution.J Biol Chem. 2014 Jun 20;289(25):17634-46. doi: 10.1074/jbc.M114.552794. Epub 2014 Apr 25. J Biol Chem. 2014. PMID: 24770420 Free PMC article.
-
RPA and RAD51: fork reversal, fork protection, and genome stability.Nat Struct Mol Biol. 2018 Jun;25(6):446-453. doi: 10.1038/s41594-018-0075-z. Epub 2018 May 28. Nat Struct Mol Biol. 2018. PMID: 29807999 Free PMC article. Review.
-
The mechanism of action of the SSB interactome reveals it is the first OB-fold family of genome guardians in prokaryotes.Protein Sci. 2021 Sep;30(9):1757-1775. doi: 10.1002/pro.4140. Epub 2021 Jun 14. Protein Sci. 2021. PMID: 34089559 Free PMC article. Review.
References
-
- Lohman T. M. & Bjornson K. P. Mechanisms of helicase-catalyzed DNA unwinding. Annu. Rev. Biochem. 65, 169–214 (1996). - PubMed
-
- Matson S. W., Bean D. W. & George J. W. DNA helicases: enzymes with essential roles in all aspects of DNA metabolism. Bioessays 16, 13–22 (1994). - PubMed
-
- Nelson S. W. & Benkovic S. J. The T4 phage UvsW protein contains both DNA unwinding and strand annealing activities. J. Biol. Chem. 282, 407–416 (2007). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
