

Recruitment into stress granules prevents irreversible aggregation of FUS protein mislocalized to the cytoplasm
- PMID: 24013423
- PMCID: PMC3865015
- DOI: 10.4161/cc.26241
Recruitment into stress granules prevents irreversible aggregation of FUS protein mislocalized to the cytoplasm
Abstract
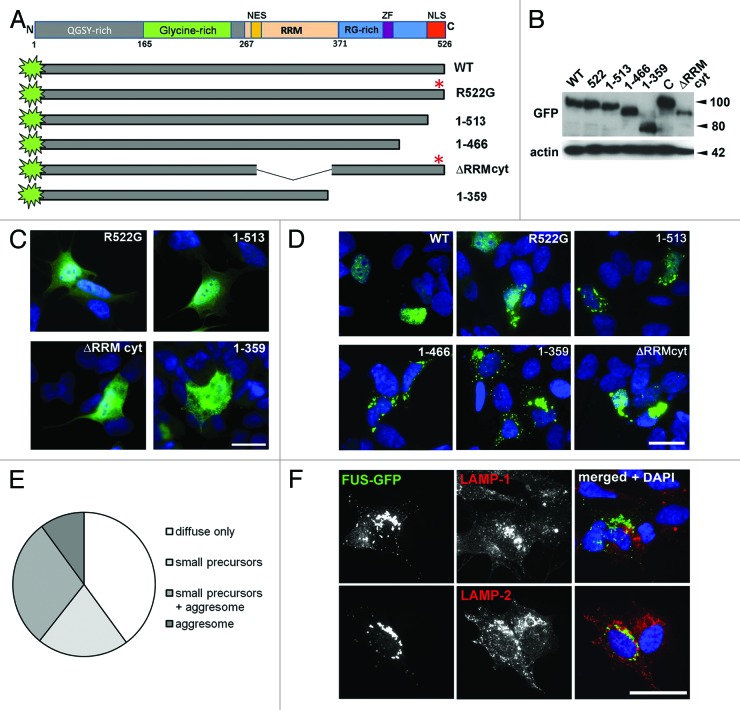
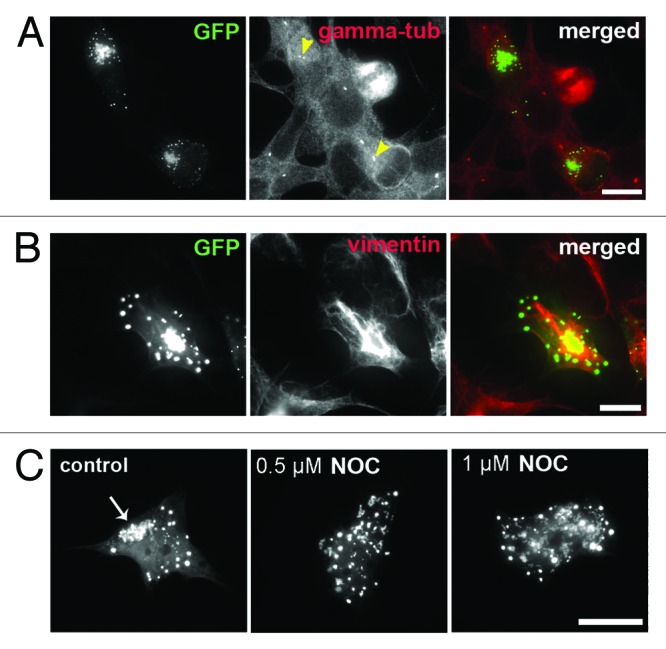
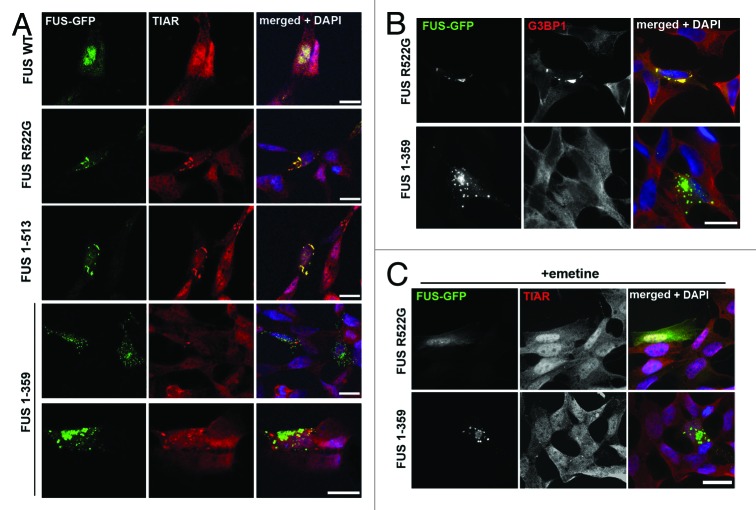
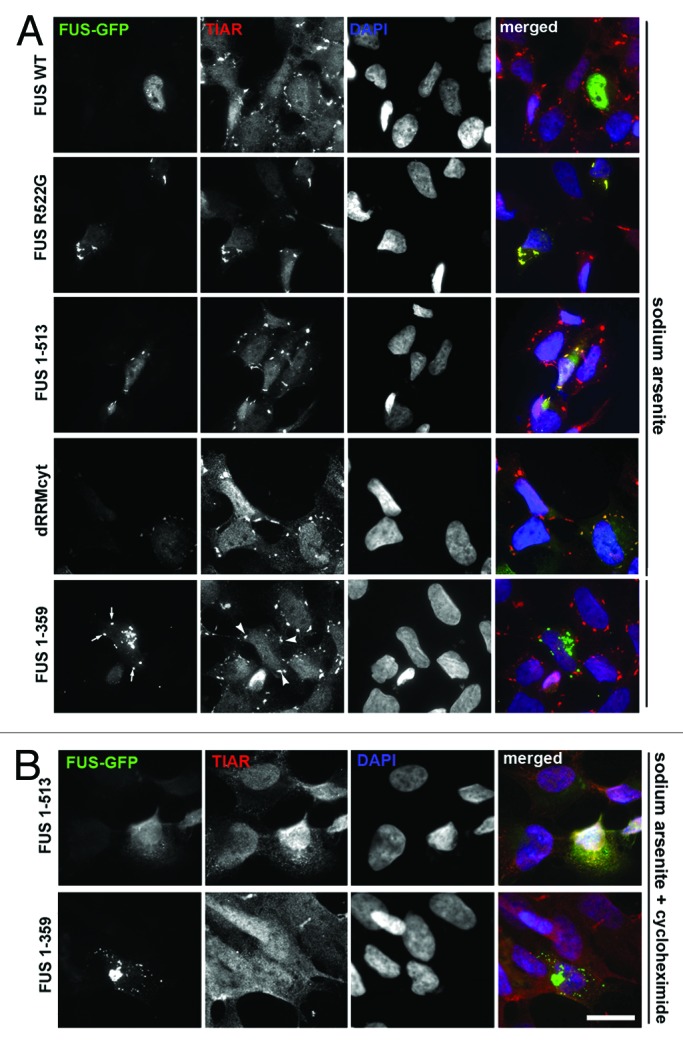
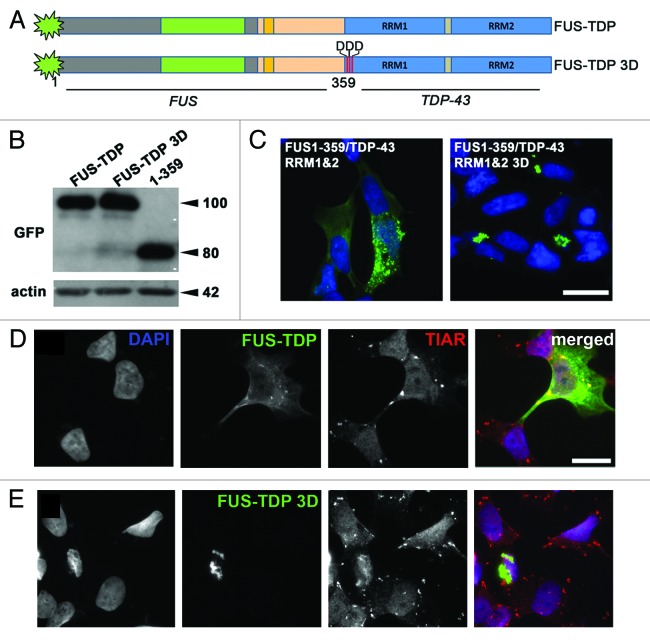
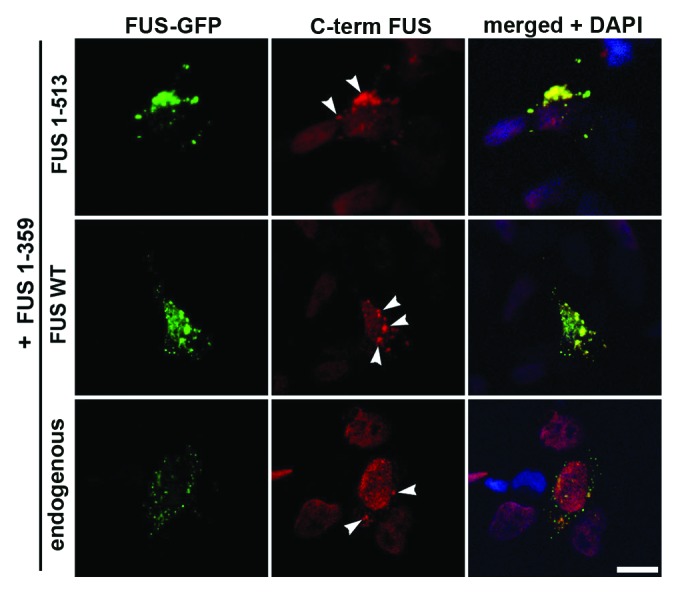
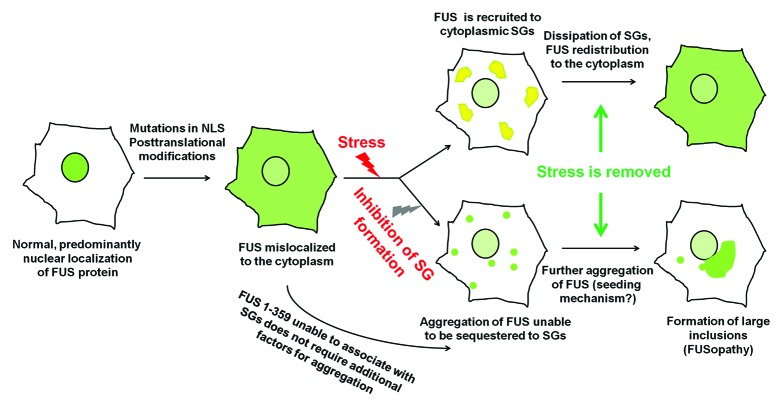
Fused in sarcoma (FUS) belongs to the group of RNA-binding proteins implicated as underlying factors in amyotrophic lateral sclerosis (ALS) and certain other neurodegenerative diseases. Multiple FUS gene mutations have been linked to hereditary forms, and aggregation of FUS protein is believed to play an important role in pathogenesis of these diseases. In cultured cells, FUS variants with disease-associated amino acid substitutions or short deletions affecting nuclear localization signal (NLS) and causing cytoplasmic mislocalization can be sequestered into stress granules (SGs). We demonstrated that disruption of motifs responsible for RNA recognition and binding not only prevents SG recruitment, but also dramatically increases the protein propensity to aggregate in the cell cytoplasm with formation of juxtanuclear structures displaying typical features of aggresomes. Functional RNA-binding domains from TAR DNA-binding protein of 43 kDa (TDP-43) fused to highly aggregation-prone C-terminally truncated FUS protein restored the ability to enter SGs and prevented aggregation of the chimeric protein. Truncated FUS was also able to trap endogenous FUS molecules in the cytoplasmic aggregates. Our data indicate that RNA binding and recruitment to SGs protect cytoplasmic FUS from aggregation, and loss of this protection may trigger its pathological aggregation in vivo.
Keywords: FUSopathy; RNA-binding proteins; amyotrophic lateral sclerosis; cytoplasmic RNP complexes; protein aggregation; proteinopathy.
Figures
Similar articles
-
Multistep process of FUS aggregation in the cell cytoplasm involves RNA-dependent and RNA-independent mechanisms.Hum Mol Genet. 2014 Oct 1;23(19):5211-26. doi: 10.1093/hmg/ddu243. Epub 2014 May 19. Hum Mol Genet. 2014. PMID: 24842888 Free PMC article.
-
Aggregation of ALS-linked FUS mutant sequesters RNA binding proteins and impairs RNA granules formation.Biochem Biophys Res Commun. 2014 Sep 26;452(3):600-7. doi: 10.1016/j.bbrc.2014.08.115. Epub 2014 Aug 28. Biochem Biophys Res Commun. 2014. PMID: 25173930
-
Stress granules in neurodegeneration--lessons learnt from TAR DNA binding protein of 43 kDa and fused in sarcoma.FEBS J. 2013 Sep;280(18):4348-70. doi: 10.1111/febs.12287. Epub 2013 May 9. FEBS J. 2013. PMID: 23587065 Review.
-
RNA-binding ability of FUS regulates neurodegeneration, cytoplasmic mislocalization and incorporation into stress granules associated with FUS carrying ALS-linked mutations.Hum Mol Genet. 2013 Mar 15;22(6):1193-205. doi: 10.1093/hmg/dds526. Epub 2012 Dec 20. Hum Mol Genet. 2013. PMID: 23257289 Free PMC article.
-
"STRESSED OUT": The role of FUS and TDP-43 in amyotrophic lateral sclerosis.Int J Biochem Cell Biol. 2020 Sep;126:105821. doi: 10.1016/j.biocel.2020.105821. Epub 2020 Aug 3. Int J Biochem Cell Biol. 2020. PMID: 32758633 Review.
Cited by
-
Phenotypic Suppression of ALS/FTD-Associated Neurodegeneration Highlights Mechanisms of Dysfunction.J Neurosci. 2019 Oct 16;39(42):8217-8224. doi: 10.1523/JNEUROSCI.1159-19.2019. J Neurosci. 2019. PMID: 31619490 Free PMC article. Review.
-
Arsenite-activated JNK signaling enhances CPEB4-Vinexin interaction to facilitate stress granule assembly and cell survival.PLoS One. 2014 Sep 19;9(9):e107961. doi: 10.1371/journal.pone.0107961. eCollection 2014. PLoS One. 2014. PMID: 25237887 Free PMC article.
-
Calcium-responsive transactivator (CREST) protein shares a set of structural and functional traits with other proteins associated with amyotrophic lateral sclerosis.Mol Neurodegener. 2015 Apr 10;10:20. doi: 10.1186/s13024-015-0014-y. Mol Neurodegener. 2015. PMID: 25888396 Free PMC article.
-
Trends in Understanding the Pathological Roles of TDP-43 and FUS Proteins.Adv Exp Med Biol. 2021;1281:243-267. doi: 10.1007/978-3-030-51140-1_15. Adv Exp Med Biol. 2021. PMID: 33433879
-
Multistep process of FUS aggregation in the cell cytoplasm involves RNA-dependent and RNA-independent mechanisms.Hum Mol Genet. 2014 Oct 1;23(19):5211-26. doi: 10.1093/hmg/ddu243. Epub 2014 May 19. Hum Mol Genet. 2014. PMID: 24842888 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous