

Regulation of constitutive and alternative splicing by PRMT5 reveals a role for Mdm4 pre-mRNA in sensing defects in the spliceosomal machinery
- PMID: 24013503
- PMCID: PMC3778243
- DOI: 10.1101/gad.219899.113
Regulation of constitutive and alternative splicing by PRMT5 reveals a role for Mdm4 pre-mRNA in sensing defects in the spliceosomal machinery
Abstract
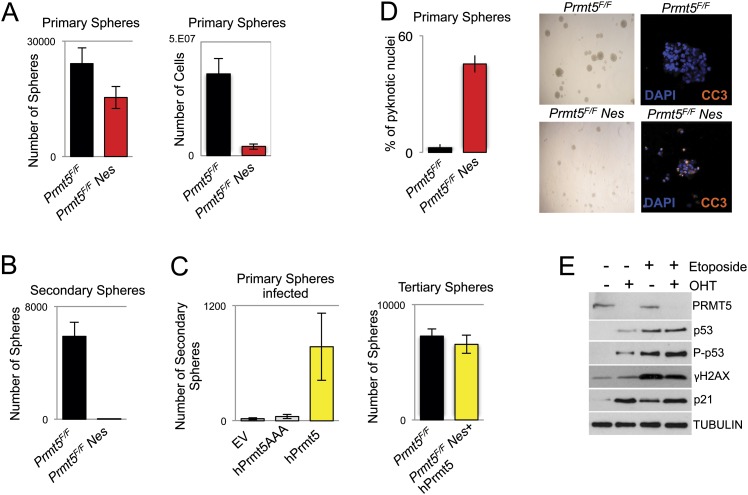
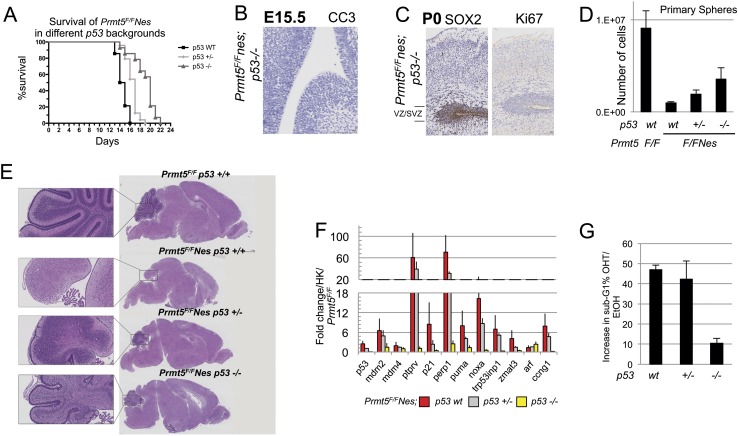
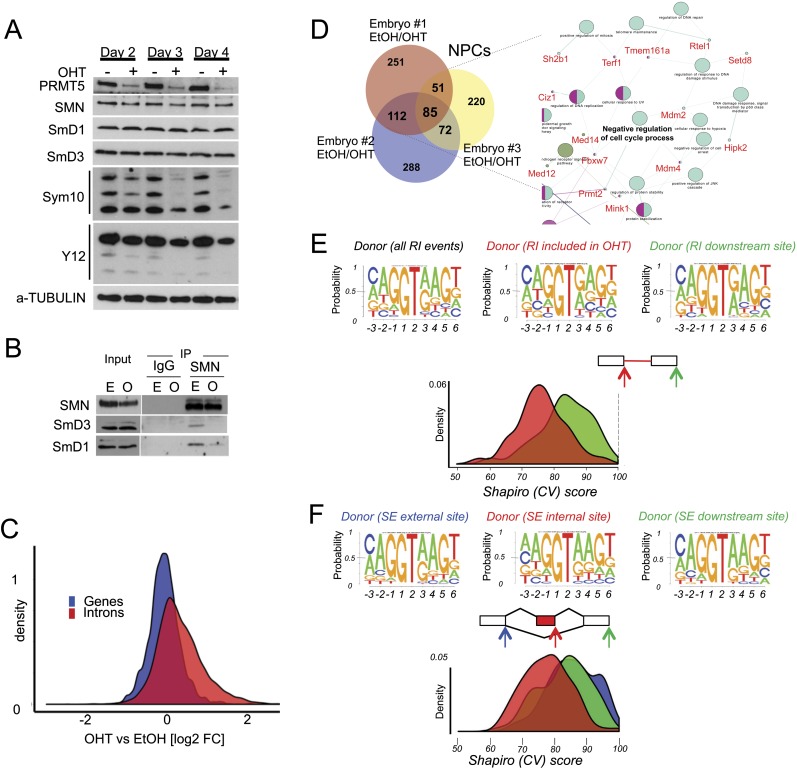
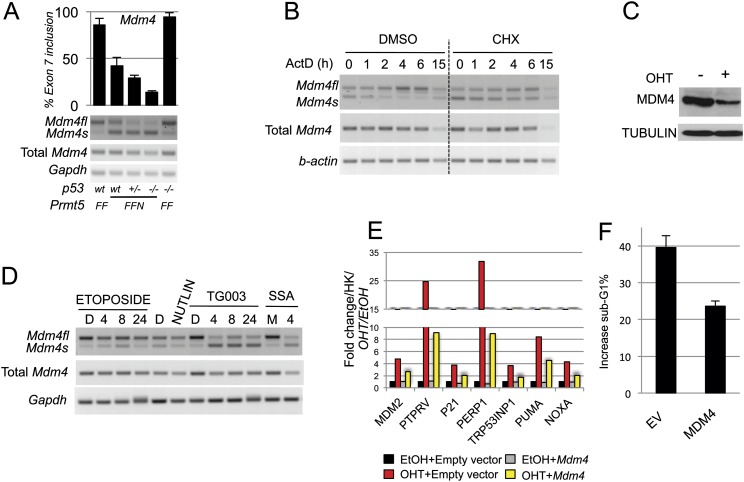
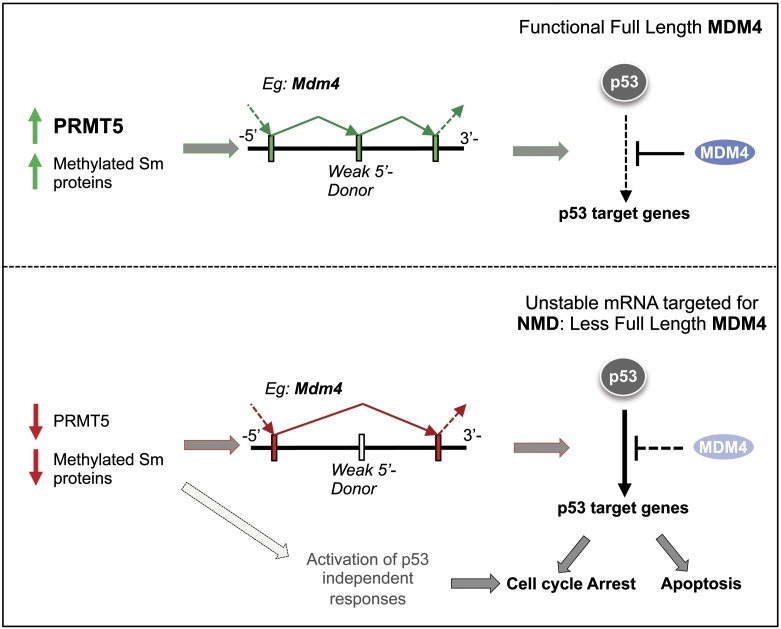
The tight control of gene expression at the level of both transcription and post-transcriptional RNA processing is essential for mammalian development. We here investigate the role of protein arginine methyltransferase 5 (PRMT5), a putative splicing regulator and transcriptional cofactor, in mammalian development. We demonstrate that selective deletion of PRMT5 in neural stem/progenitor cells (NPCs) leads to postnatal death in mice. At the molecular level, the absence of PRMT5 results in reduced methylation of Sm proteins, aberrant constitutive splicing, and the alternative splicing of specific mRNAs with weak 5' donor sites. Intriguingly, the products of these mRNAs are, among others, several proteins regulating cell cycle progression. We identify Mdm4 as one of these key mRNAs that senses the defects in the spliceosomal machinery and transduces the signal to activate the p53 response, providing a mechanistic explanation of the phenotype observed in vivo. Our data demonstrate that PRMT5 is a master regulator of splicing in mammals and uncover a new role for the Mdm4 pre-mRNA, which could be exploited for anti-cancer therapy.
Keywords: MDM4; PRMT5; arginine methylation; development; p53; splicing.
Figures


Similar articles
-
A methyl transferase links the circadian clock to the regulation of alternative splicing.Nature. 2010 Nov 4;468(7320):112-6. doi: 10.1038/nature09470. Epub 2010 Oct 20. Nature. 2010. PMID: 20962777
-
Deletion of RBMX RGG/RG motif in Shashi-XLID syndrome leads to aberrant p53 activation and neuronal differentiation defects.Cell Rep. 2021 Jul 13;36(2):109337. doi: 10.1016/j.celrep.2021.109337. Cell Rep. 2021. PMID: 34260915
-
Activation of the p53-MDM4 regulatory axis defines the anti-tumour response to PRMT5 inhibition through its role in regulating cellular splicing.Sci Rep. 2018 Jun 26;8(1):9711. doi: 10.1038/s41598-018-28002-y. Sci Rep. 2018. PMID: 29946150 Free PMC article.
-
Alternative splicing at the right time.RNA Biol. 2011 Nov-Dec;8(6):954-9. doi: 10.4161/rna.8.6.17336. Epub 2011 Nov 1. RNA Biol. 2011. PMID: 21941124 Free PMC article. Review.
-
Splicing to Keep Cycling: The Importance of Pre-mRNA Splicing during the Cell Cycle.Trends Genet. 2021 Mar;37(3):266-278. doi: 10.1016/j.tig.2020.08.013. Epub 2020 Sep 16. Trends Genet. 2021. PMID: 32950269 Review.
Cited by
-
Histone H2A and H4 N-terminal tails are positioned by the MEP50 WD repeat protein for efficient methylation by the PRMT5 arginine methyltransferase.J Biol Chem. 2015 Apr 10;290(15):9674-89. doi: 10.1074/jbc.M115.636894. Epub 2015 Feb 24. J Biol Chem. 2015. PMID: 25713080 Free PMC article.
-
MYCN and SNRPD3 cooperate to maintain a balance of alternative splicing events that drives neuroblastoma progression.Oncogene. 2024 Jan;43(5):363-377. doi: 10.1038/s41388-023-02897-y. Epub 2023 Dec 4. Oncogene. 2024. PMID: 38049564 Free PMC article.
-
Recruitment of the NineTeen Complex to the activated spliceosome requires AtPRMT5.Proc Natl Acad Sci U S A. 2016 May 10;113(19):5447-52. doi: 10.1073/pnas.1522458113. Epub 2016 Apr 25. Proc Natl Acad Sci U S A. 2016. PMID: 27114555 Free PMC article.
-
Loss-of-function mutation in PRMT9 causes abnormal synapse development by dysregulation of RNA alternative splicing.Nat Commun. 2024 Apr 1;15(1):2809. doi: 10.1038/s41467-024-47107-9. Nat Commun. 2024. PMID: 38561334 Free PMC article.
-
TDRD6 mediates early steps of spliceosome maturation in primary spermatocytes.PLoS Genet. 2017 Mar 6;13(3):e1006660. doi: 10.1371/journal.pgen.1006660. eCollection 2017 Mar. PLoS Genet. 2017. PMID: 28263986 Free PMC article.
References
-
- Allende-Vega N, Dayal S, Agarwala U, Sparks A, Bourdon JC, Saville MK 2013. p53 is activated in response to disruption of the pre-mRNA splicing machinery. Oncogene 32: 1–14 - PubMed
-
- Barbosa-Morais NL, Irimia M, Pan Q, Xiong HY, Gueroussov S, Lee LJ, Slobodeniuc V, Kutter C, Watt S, Colak R, et al. 2012. The evolutionary landscape of alternative splicing in vertebrate species. Science 338: 1587–1593 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous