

Mutually exclusive signaling signatures define the hepatic and pancreatic progenitor cell lineage divergence
- PMID: 24013505
- PMCID: PMC3778245
- DOI: 10.1101/gad.220244.113
Mutually exclusive signaling signatures define the hepatic and pancreatic progenitor cell lineage divergence
Abstract
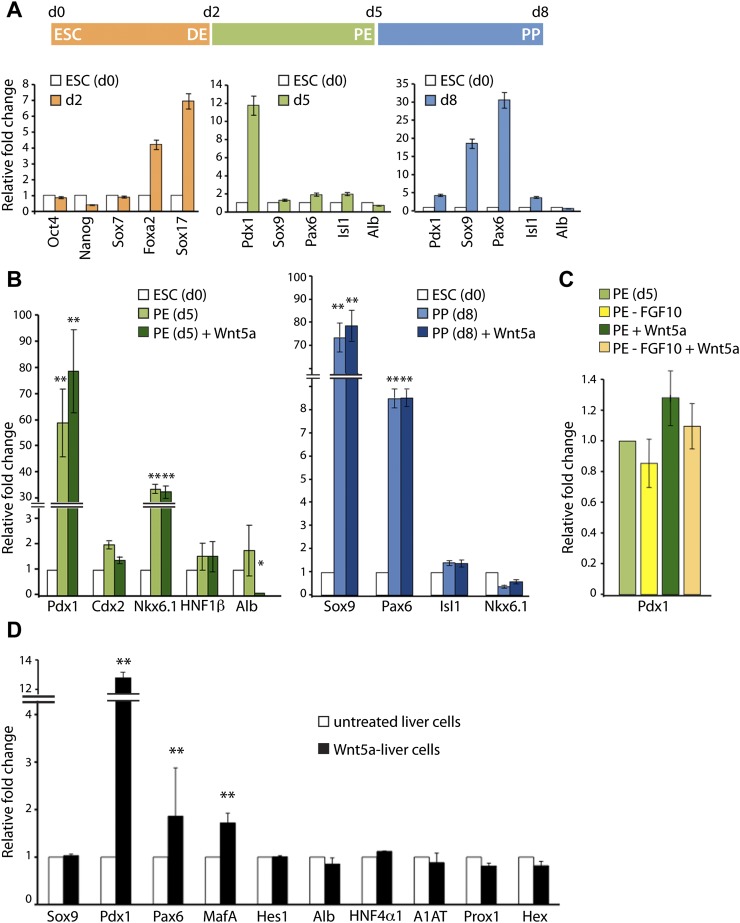
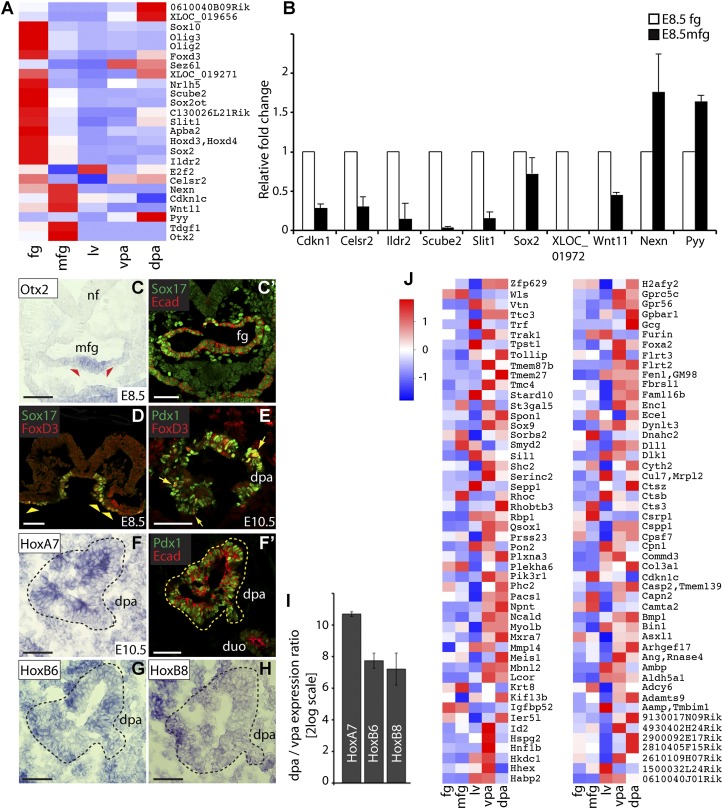
Understanding how distinct cell types arise from multipotent progenitor cells is a major quest in stem cell biology. The liver and pancreas share many aspects of their early development and possibly originate from a common progenitor. However, how liver and pancreas cells diverge from a common endoderm progenitor population and adopt specific fates remains elusive. Using RNA sequencing (RNA-seq), we defined the molecular identity of liver and pancreas progenitors that were isolated from the mouse embryo at two time points, spanning the period when the lineage decision is made. The integration of temporal and spatial gene expression profiles unveiled mutually exclusive signaling signatures in hepatic and pancreatic progenitors. Importantly, we identified the noncanonical Wnt pathway as a potential developmental regulator of this fate decision and capable of inducing the pancreas program in endoderm and liver cells. Our study offers an unprecedented view of gene expression programs in liver and pancreas progenitors and forms the basis for formulating lineage-reprogramming strategies to convert adult hepatic cells into pancreatic cells.
Keywords: RNA-seq; Wnt signaling; liver; mouse; pancreas.
Figures
Similar articles
-
Quantitative lineage analysis identifies a hepato-pancreato-biliary progenitor niche.Nature. 2021 Sep;597(7874):87-91. doi: 10.1038/s41586-021-03844-1. Epub 2021 Aug 25. Nature. 2021. PMID: 34433966
-
Hyperglycemia impedes definitive endoderm differentiation of human embryonic stem cells by modulating histone methylation patterns.Cell Tissue Res. 2017 Jun;368(3):563-578. doi: 10.1007/s00441-017-2583-2. Epub 2017 Mar 10. Cell Tissue Res. 2017. PMID: 28283910
-
Ontogenesis of hepatic and pancreatic stem cells.Stem Cell Rev Rep. 2012 Jun;8(2):586-96. doi: 10.1007/s12015-012-9350-2. Stem Cell Rev Rep. 2012. PMID: 22350509 Review.
-
A single-cell-resolution fate map of endoderm reveals demarcation of pancreatic progenitors by cell cycle.Proc Natl Acad Sci U S A. 2021 Jun 22;118(25):e2025793118. doi: 10.1073/pnas.2025793118. Proc Natl Acad Sci U S A. 2021. PMID: 34161274 Free PMC article.
-
Pioneer factors, genetic competence, and inductive signaling: programming liver and pancreas progenitors from the endoderm.Cold Spring Harb Symp Quant Biol. 2008;73:119-26. doi: 10.1101/sqb.2008.73.040. Epub 2008 Nov 21. Cold Spring Harb Symp Quant Biol. 2008. PMID: 19028990 Free PMC article. Review.
Cited by
-
Vascularized Organoids: A More Complete Model.Int J Stem Cells. 2021 May 30;14(2):127-137. doi: 10.15283/ijsc20143. Int J Stem Cells. 2021. PMID: 33377457 Free PMC article. Review.
-
Quantitative lineage analysis identifies a hepato-pancreato-biliary progenitor niche.Nature. 2021 Sep;597(7874):87-91. doi: 10.1038/s41586-021-03844-1. Epub 2021 Aug 25. Nature. 2021. PMID: 34433966
-
Modeling HNF1B-associated monogenic diabetes using human iPSCs reveals an early stage impairment of the pancreatic developmental program.Stem Cell Reports. 2021 Sep 14;16(9):2289-2304. doi: 10.1016/j.stemcr.2021.07.018. Epub 2021 Aug 26. Stem Cell Reports. 2021. PMID: 34450036 Free PMC article.
-
MicroRNA-7 regulates endocrine progenitor delamination and endocrine cell mass in developing pancreatic islets.iScience. 2024 Jun 20;27(7):110332. doi: 10.1016/j.isci.2024.110332. eCollection 2024 Jul 19. iScience. 2024. PMID: 39055950 Free PMC article.
-
Chromatin features, RNA polymerase II and the comparative expression of lens genes encoding crystallins, transcription factors, and autophagy mediators.Mol Vis. 2015 Aug 28;21:955-73. eCollection 2015. Mol Vis. 2015. PMID: 26330747 Free PMC article.
References
-
- Alexa A, Rahnenführer J, Lengauer T 2006. Improved scoring of functional groups from gene expression data by decorrelating GO graph structure. Bioinformatics 22: 1600–1607 - PubMed
-
- Angers S, Moon R 2009. Proximal events in Wnt signal transduction. Nat Rev Mol Cell Biol 10: 468–477 - PubMed
-
- Burke Z, Oliver G 2002. Prox1 is an early specific marker for the developing liver and pancreas in the mammalian foregut endoderm. Mech Dev 118: 147–155 - PubMed
-
- Chalmers A, Slack J, Beck C 2000. Regional gene expression in the epithelia of the Xenopus tadpole gut. Mech Dev 96: 125–128 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases