

Activation of the Ca²+-sensing receptor induces deposition of tight junction components to the epithelial cell plasma membrane
- PMID: 24013548
- PMCID: PMC3828589
- DOI: 10.1242/jcs.127555
Activation of the Ca²+-sensing receptor induces deposition of tight junction components to the epithelial cell plasma membrane
Abstract
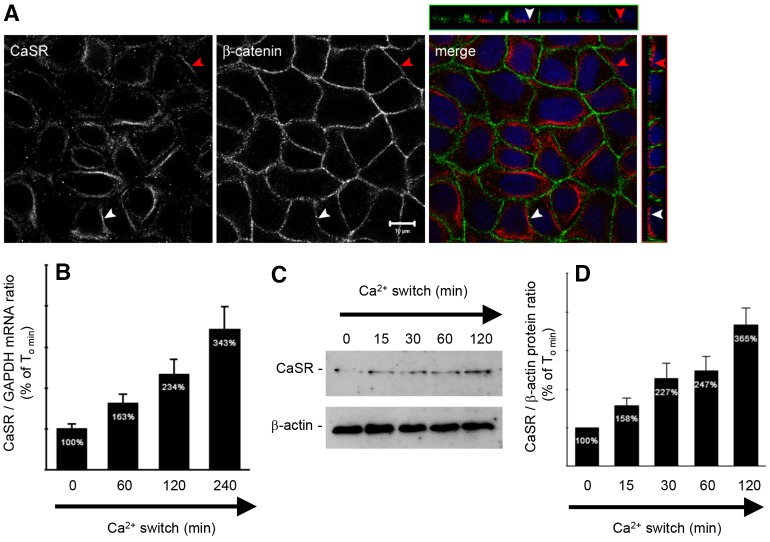
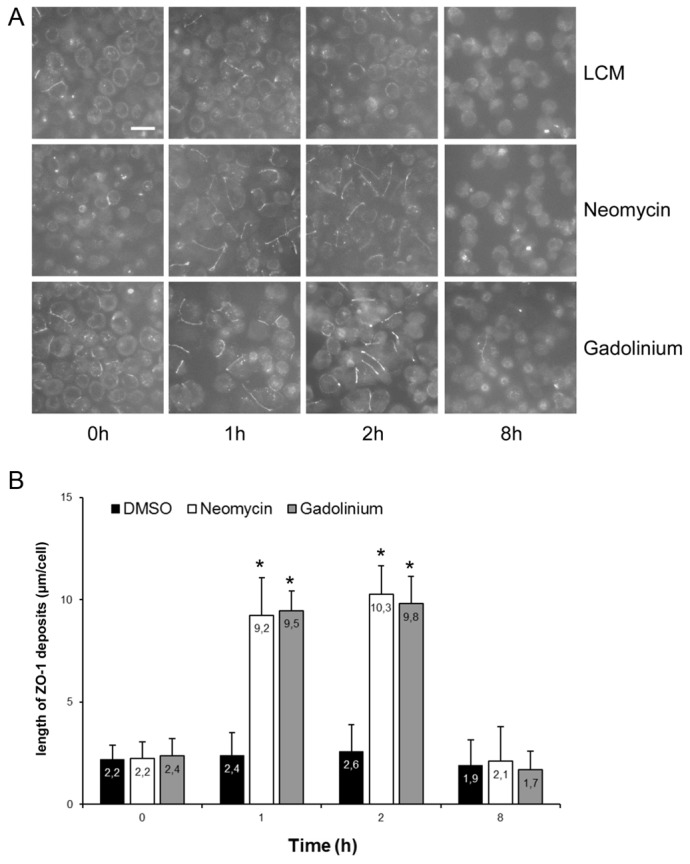
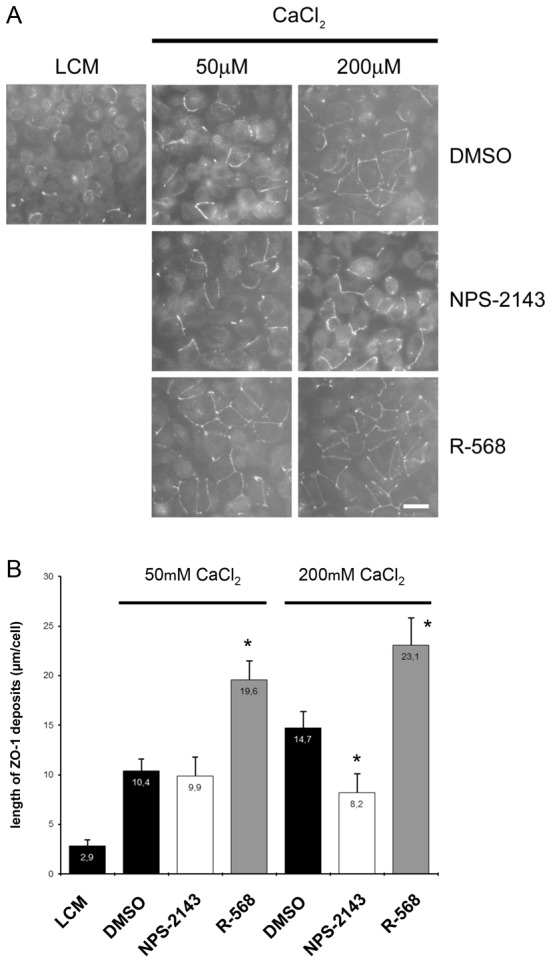
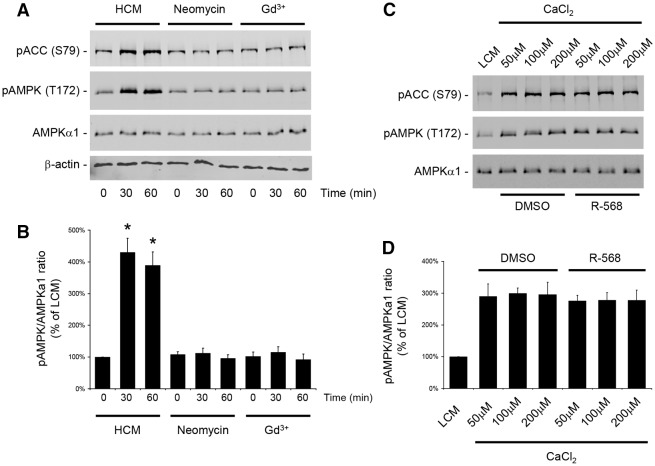
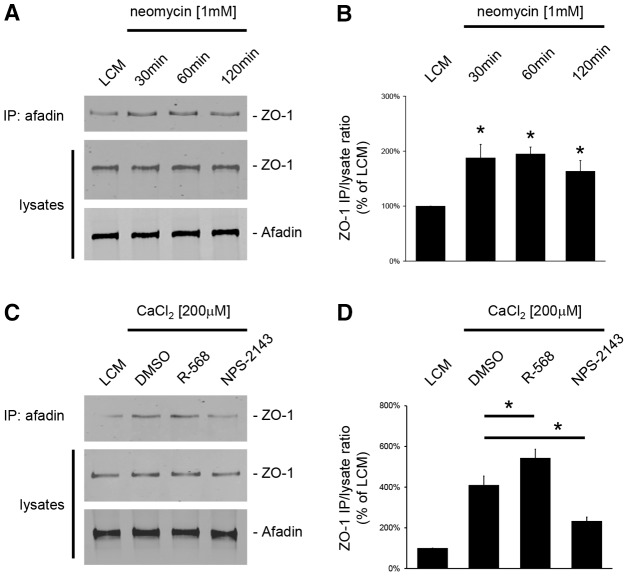
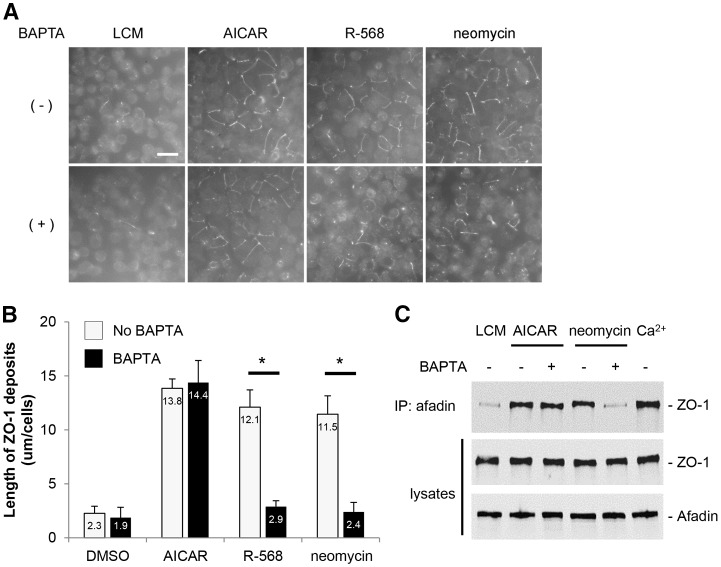
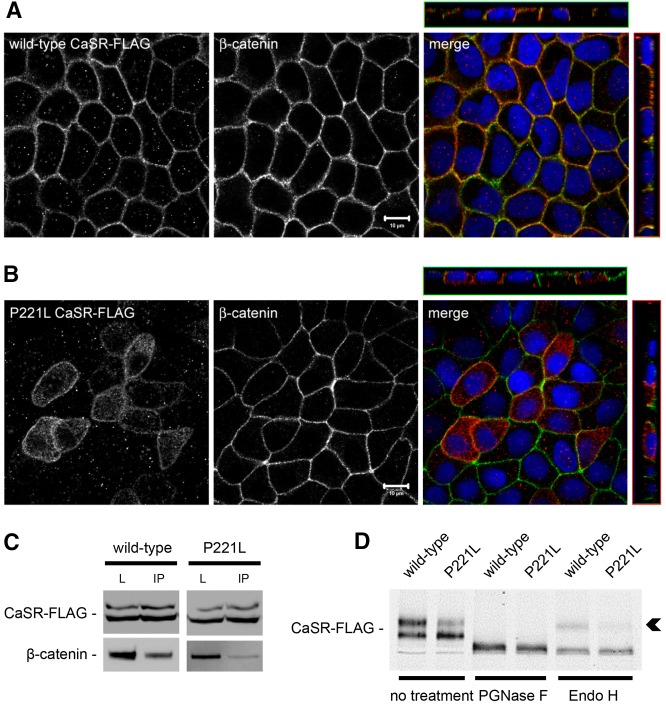
The Ca(2+)-sensing receptor (CaSR) belongs to the G-protein-coupled receptor superfamily and plays essential roles in divalent ion homeostasis and cell differentiation. Because extracellular Ca(2+) is essential for the development of stable epithelial tight junctions (TJs), we hypothesized that the CaSR participates in regulating TJ assembly. We first assessed the expression of the CaSR in Madin-Darby canine kidney (MDCK) cells at steady state and following manipulations that modulate TJ assembly. Next, we examined the effects of CaSR agonists and antagonists on TJ assembly. Immunofluorescence studies indicate that endogenous CaSR is located at the basolateral pole of MDCK cells. Stable transfection of human CaSR in MDCK cells further reveals that this protein co-distributes with β-catenin on the basolateral membrane. Switching MDCK cells from low-Ca(2+) medium to medium containing a normal Ca(2+) concentration significantly increases CaSR expression at both the mRNA and protein levels. Exposure of MDCK cells maintained in low-Ca(2+) conditions to the CaSR agonists neomycin, Gd(3+) or R-568 causes the transient relocation of the tight junction components ZO-1 and occludin to sites of cell-cell contact, while inducing no significant changes in the expression of mRNAs encoding junction-associated proteins. Stimulation of CaSR also increases the interaction between ZO-1 and the F-actin-binding protein I-afadin. This effect does not involve activation of the AMP-activated protein kinase. By contrast, CaSR inhibition by NPS-2143 significantly decreases interaction of ZO-1 with I-afadin and reduces deposition of ZO-1 at the cell surface following a Ca(2+) switch from 5 µM to 200 µM [Ca(2+)]e. Pre-exposure of MDCK cells to the cell-permeant Ca(2+) chelator BAPTA-AM, similarly prevents TJ assembly caused by CaSR activation. Finally, stable transfection of MDCK cells with a cDNA encoding a human disease-associated gain-of-function mutant form of the CaSR increases the transepithelial electrical resistance of these cells in comparison to expression of the wild-type human CaSR. These observations suggest that the CaSR participates in regulating TJ assembly.
Keywords: Calcium-sensing receptor; Epithelia; Tight junction.
Figures
References
-
- Balda M. S., González-Mariscal L., Contreras R. G., Macias-Silva M., Torres-Marquez M. E., García-Sáinz J. A., Cereijido M. (1991). Assembly and sealing of tight junctions: possible participation of G-proteins, phospholipase C, protein kinase C and calmodulin. J. Membr. Biol. 122, 193–202 10.1007/BF01871420 - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
