

The extraordinary evolutionary history of the reticuloendotheliosis viruses
- PMID: 24013706
- PMCID: PMC3754887
- DOI: 10.1371/journal.pbio.1001642
The extraordinary evolutionary history of the reticuloendotheliosis viruses
Abstract
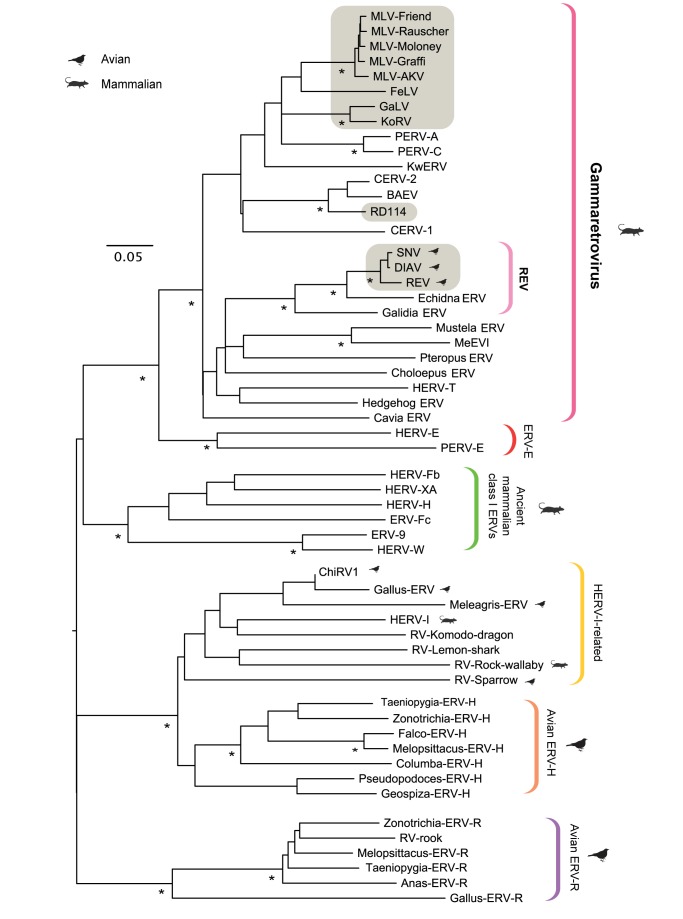
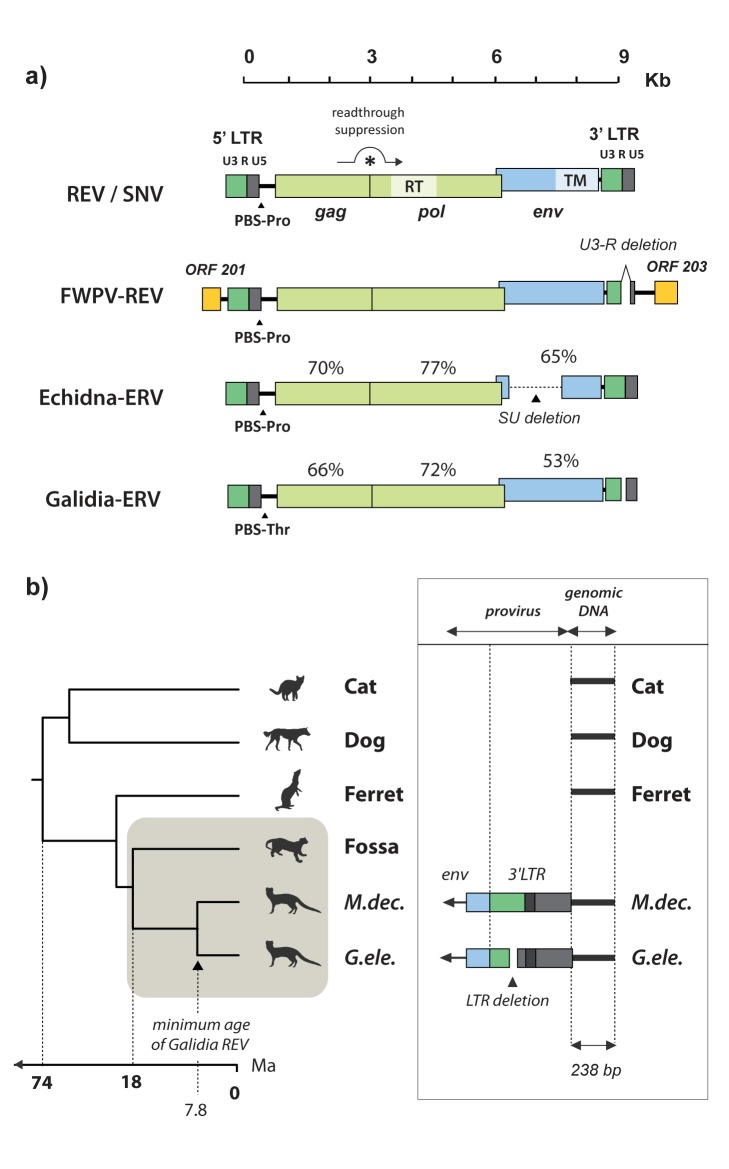
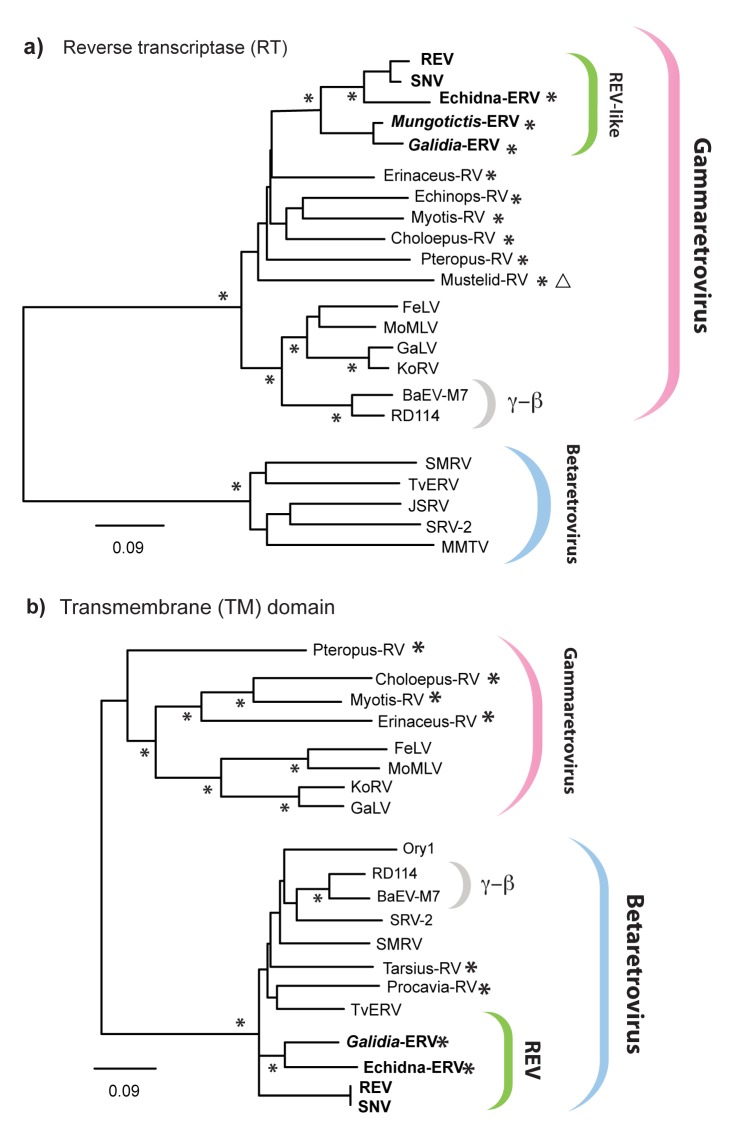
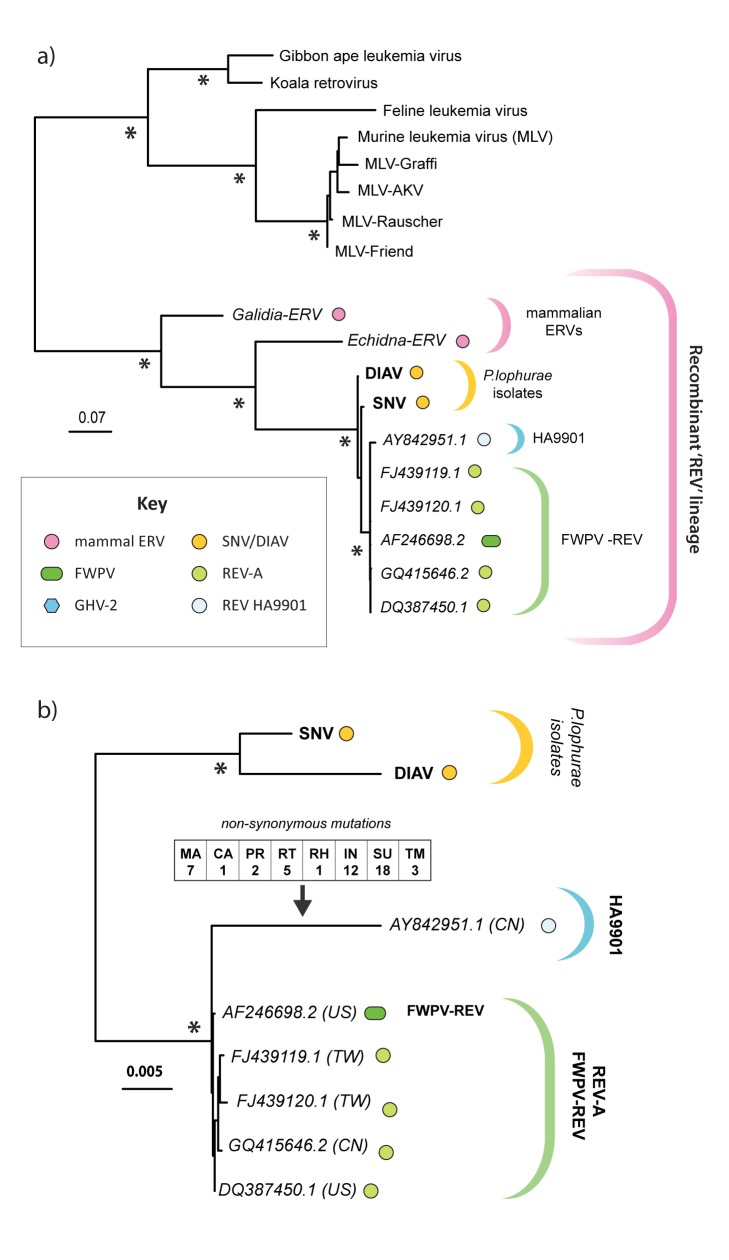
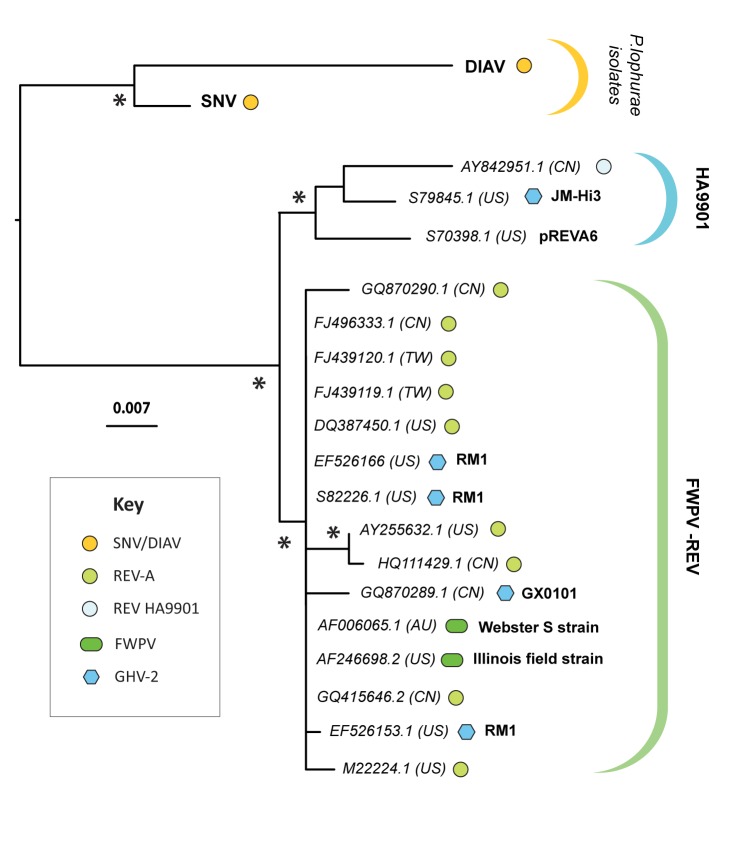
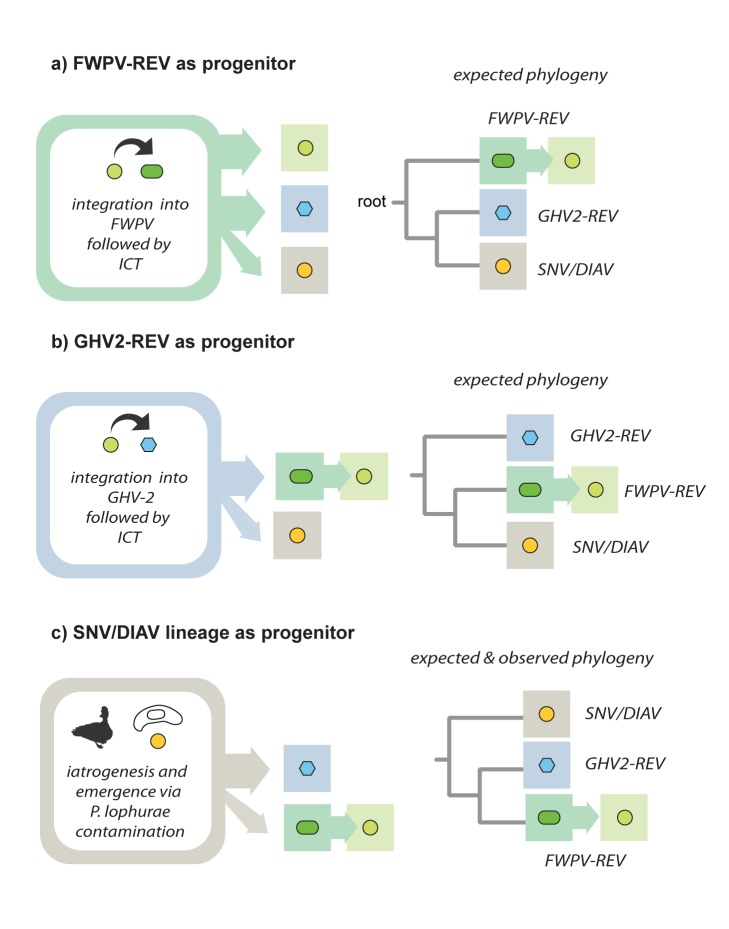
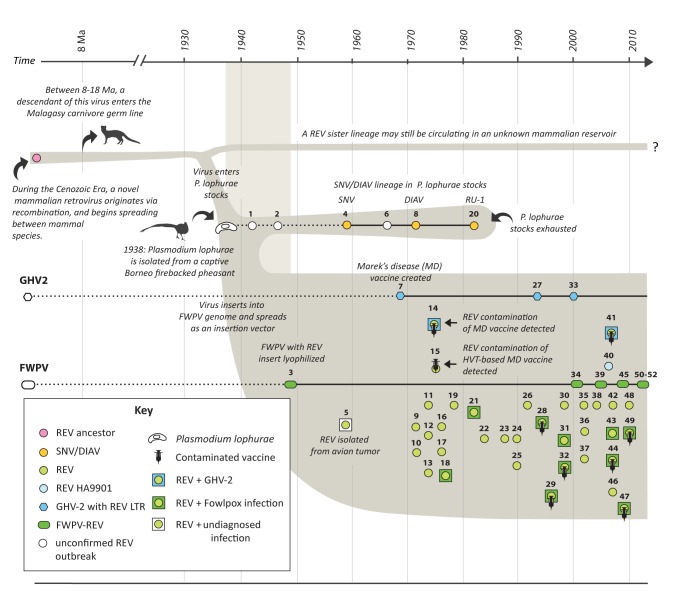
The reticuloendotheliosis viruses (REVs) comprise several closely related amphotropic retroviruses isolated from birds. These viruses exhibit several highly unusual characteristics that have not so far been adequately explained, including their extremely close relationship to mammalian retroviruses, and their presence as endogenous sequences within the genomes of certain large DNA viruses. We present evidence for an iatrogenic origin of REVs that accounts for these phenomena. Firstly, we identify endogenous retroviral fossils in mammalian genomes that share a unique recombinant structure with REVs-unequivocally demonstrating that REVs derive directly from mammalian retroviruses. Secondly, through sequencing of archived REV isolates, we confirm that contaminated Plasmodium lophurae stocks have been the source of multiple REV outbreaks in experimentally infected birds. Finally, we show that both phylogenetic and historical evidence support a scenario wherein REVs originated as mammalian retroviruses that were accidentally introduced into avian hosts in the late 1930s, during experimental studies of P. lophurae, and subsequently integrated into the fowlpox virus (FWPV) and gallid herpesvirus type 2 (GHV-2) genomes, generating recombinant DNA viruses that now circulate in wild birds and poultry. Our findings provide a novel perspective on the origin and evolution of REV, and indicate that horizontal gene transfer between virus families can expand the impact of iatrogenic transmission events.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
First Complete Genome of Reticuloendotheliosis Virus in a Mallard Duck from Brazil: Phylogenetic Insights and Evolutionary Analysis.Pathogens. 2025 Feb 13;14(2):189. doi: 10.3390/pathogens14020189. Pathogens. 2025. PMID: 40005564 Free PMC article.
-
Detection of Fowlpox virus carrying distinct genome segments of Reticuloendotheliosis virus.Virus Res. 2019 Jan 15;260:53-59. doi: 10.1016/j.virusres.2018.10.017. Epub 2018 Oct 22. Virus Res. 2019. PMID: 30359622
-
Emergence of reticuloendotheliosis virus in pigeons in Guangdong Province, Southern China.Arch Virol. 2016 Jul;161(7):2007-11. doi: 10.1007/s00705-016-2870-3. Epub 2016 Apr 27. Arch Virol. 2016. PMID: 27120185
-
Reticuloendotheliosis viruses and derived vectors for human gene therapy.Front Biosci. 2003 May 1;8:d801-17. doi: 10.2741/955. Front Biosci. 2003. PMID: 12700113 Review.
-
Transformation of avian lymphoid cells by reticuloendotheliosis virus.Mutat Res. 1988 Jan;195(1):79-90. doi: 10.1016/0165-1110(88)90016-4. Mutat Res. 1988. PMID: 2827014 Review.
Cited by
-
Co-option of endogenous viral sequences for host cell function.Curr Opin Virol. 2017 Aug;25:81-89. doi: 10.1016/j.coviro.2017.07.021. Epub 2017 Aug 16. Curr Opin Virol. 2017. PMID: 28818736 Free PMC article. Review.
-
Viruses' evolvement as a never stopping perpetuum mobile.J Virol Methods. 2021 Mar;289:114037. doi: 10.1016/j.jviromet.2020.114037. Epub 2020 Dec 5. J Virol Methods. 2021. PMID: 33290787 Free PMC article. Review. No abstract available.
-
First Complete Genome of Reticuloendotheliosis Virus in a Mallard Duck from Brazil: Phylogenetic Insights and Evolutionary Analysis.Pathogens. 2025 Feb 13;14(2):189. doi: 10.3390/pathogens14020189. Pathogens. 2025. PMID: 40005564 Free PMC article.
-
Cross-species transmission of an ancient endogenous retrovirus and convergent co-option of its envelope gene in two mammalian orders.PLoS Genet. 2022 Oct 14;18(10):e1010458. doi: 10.1371/journal.pgen.1010458. eCollection 2022 Oct. PLoS Genet. 2022. PMID: 36240227 Free PMC article.
-
Existence of variant strains Fowlpox virus integrated with Reticuloendotheliosis virus in its genome in field isolates in Tanzania.Trop Anim Health Prod. 2014 Jun;46(5):711-6. doi: 10.1007/s11250-014-0552-2. Epub 2014 Feb 21. Trop Anim Health Prod. 2014. PMID: 24557589
References
-
- Purchase HG, Ludford C, Nazerian K, Cox HW (1973) A new group of oncogenic viruses: reticuloendotheliosis, chick syncytial, duck infectious anemia, and spleen necrosis viruses. J Natl Cancer Inst 51: 489–499. - PubMed
-
- Robinson FR, Twiehaus MJ (1974) Isolation of the avian reticuloendothelial virus (strain T). Avian Dis 18: 278–288. - PubMed
-
- Fadly AM, Zavala G, Witter RL (2011) Reticuloendotheliosis. In: Saif YM, editor. Diseases of poultry. New York: John Wiley Sons.
-
- Stoye JP (2012) Studies of endogenous retroviruses reveal a continuing evolutionary saga. Nat Rev Microbiol 10: 395–406. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
