

The CK2 kinase stabilizes CLOCK and represses its activity in the Drosophila circadian oscillator
- PMID: 24013921
- PMCID: PMC3754892
- DOI: 10.1371/journal.pbio.1001645
The CK2 kinase stabilizes CLOCK and represses its activity in the Drosophila circadian oscillator
Abstract
Phosphorylation is a pivotal regulatory mechanism for protein stability and activity in circadian clocks regardless of their evolutionary origin. It determines the speed and strength of molecular oscillations by acting on transcriptional activators and their repressors, which form negative feedback loops. In Drosophila, the CK2 kinase phosphorylates and destabilizes the PERIOD (PER) and TIMELESS (TIM) proteins, which inhibit CLOCK (CLK) transcriptional activity. Here we show that CK2 also targets the CLK activator directly. Downregulating the activity of the catalytic α subunit of CK2 induces CLK degradation, even in the absence of PER and TIM. Unexpectedly, the regulatory β subunit of the CK2 holoenzyme is not required for the regulation of CLK stability. In addition, downregulation of CK2α activity decreases CLK phosphorylation and increases per and tim transcription. These results indicate that CK2 inhibits CLK degradation while reducing its activity. Since the CK1 kinase promotes CLK degradation, we suggest that CLK stability and transcriptional activity result from counteracting effects of CK1 and CK2.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures





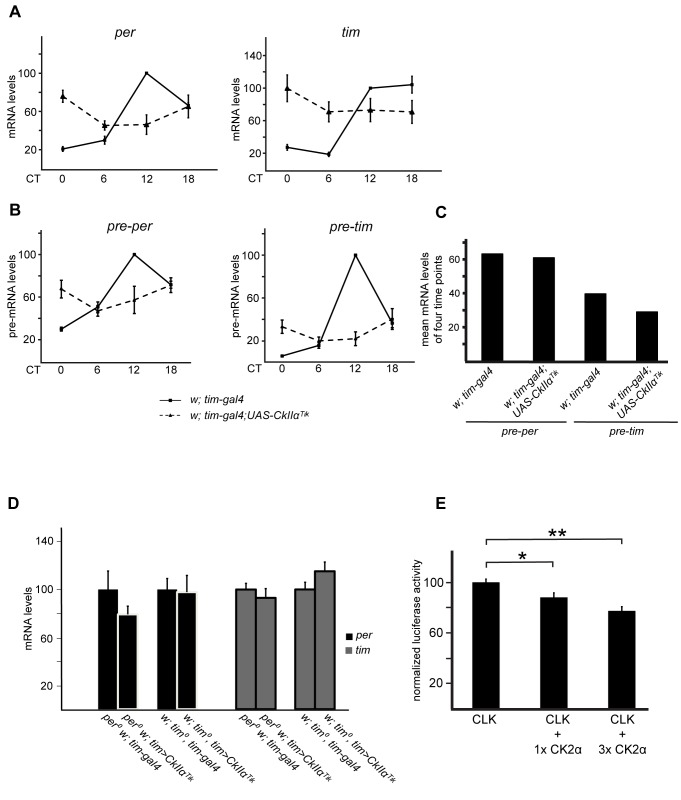
Similar articles
-
Phosphorylation of the transcription activator CLOCK regulates progression through a ∼ 24-h feedback loop to influence the circadian period in Drosophila.J Biol Chem. 2014 Jul 11;289(28):19681-93. doi: 10.1074/jbc.M114.568493. Epub 2014 May 28. J Biol Chem. 2014. PMID: 24872414 Free PMC article.
-
CLOCKWORK ORANGE promotes CLOCK-CYCLE activation via the putative Drosophila ortholog of CLOCK INTERACTING PROTEIN CIRCADIAN.Curr Biol. 2021 Oct 11;31(19):4207-4218.e4. doi: 10.1016/j.cub.2021.07.017. Epub 2021 Jul 30. Curr Biol. 2021. PMID: 34331859 Free PMC article.
-
NEMO kinase contributes to core period determination by slowing the pace of the Drosophila circadian oscillator.Curr Biol. 2011 May 10;21(9):756-61. doi: 10.1016/j.cub.2011.02.037. Epub 2011 Apr 21. Curr Biol. 2011. PMID: 21514156 Free PMC article.
-
Transcriptional feedback loop regulation, function, and ontogeny in Drosophila.Cold Spring Harb Symp Quant Biol. 2007;72:437-44. doi: 10.1101/sqb.2007.72.009. Cold Spring Harb Symp Quant Biol. 2007. PMID: 18419302 Free PMC article. Review.
-
Molecular genetic analysis of circadian timekeeping in Drosophila.Adv Genet. 2011;74:141-73. doi: 10.1016/B978-0-12-387690-4.00005-2. Adv Genet. 2011. PMID: 21924977 Free PMC article. Review.
Cited by
-
Phosphorylation of the transcription activator CLOCK regulates progression through a ∼ 24-h feedback loop to influence the circadian period in Drosophila.J Biol Chem. 2014 Jul 11;289(28):19681-93. doi: 10.1074/jbc.M114.568493. Epub 2014 May 28. J Biol Chem. 2014. PMID: 24872414 Free PMC article.
-
The Function, Regulation, and Mechanism of Protein Turnover in Circadian Systems in Neurospora and Other Species.Int J Mol Sci. 2024 Feb 22;25(5):2574. doi: 10.3390/ijms25052574. Int J Mol Sci. 2024. PMID: 38473819 Free PMC article. Review.
-
Coordination between Differentially Regulated Circadian Clocks Generates Rhythmic Behavior.Cold Spring Harb Perspect Biol. 2018 Jul 2;10(7):a033589. doi: 10.1101/cshperspect.a033589. Cold Spring Harb Perspect Biol. 2018. PMID: 28893860 Free PMC article. Review.
-
Circadian Control of Protein Synthesis.Bioessays. 2025 Mar;47(3):e202300158. doi: 10.1002/bies.202300158. Epub 2024 Dec 12. Bioessays. 2025. PMID: 39668398 Free PMC article. Review.
-
CK1α Collaborates with DOUBLETIME to Regulate PERIOD Function in the Drosophila Circadian Clock.J Neurosci. 2018 Dec 12;38(50):10631-10643. doi: 10.1523/JNEUROSCI.0871-18.2018. Epub 2018 Oct 29. J Neurosci. 2018. PMID: 30373768 Free PMC article.
References
-
- Weber F, Zorn D, Rademacher C, Hung HC (2011) Post-translational timing mechanisms of the Drosophila circadian clock. FEBS Lett 585: 1443–1449. - PubMed
-
- Kloss B, Price JL, Saez L, Blau J, Rothenfluh A, et al. (1998) The Drosophila clock gene double-time encodes a protein closely related to human casein kinase Iepsilon. Cell 94: 97–107. - PubMed
-
- Price JL, Blau J, Rothenfluh A, Abodeely M, Kloss B, et al. (1998) double-time is a novel Drosophila clock gene that regulates PERIOD protein accumulation. Cell 94: 83–95. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
