

Restoring the procofactor state of factor Va-like variants by complementation with B-domain peptides
- PMID: 24014022
- PMCID: PMC3798483
- DOI: 10.1074/jbc.M113.506840
Restoring the procofactor state of factor Va-like variants by complementation with B-domain peptides
Abstract
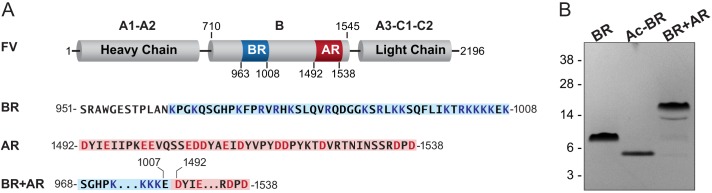
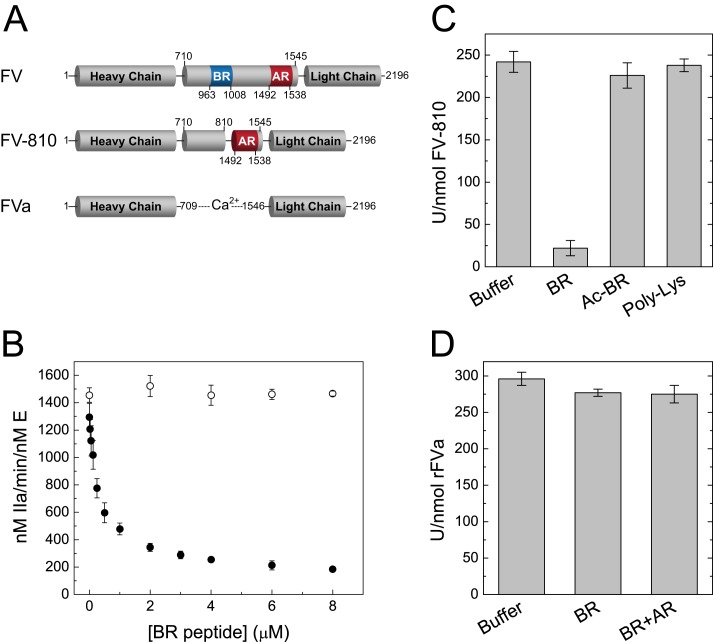
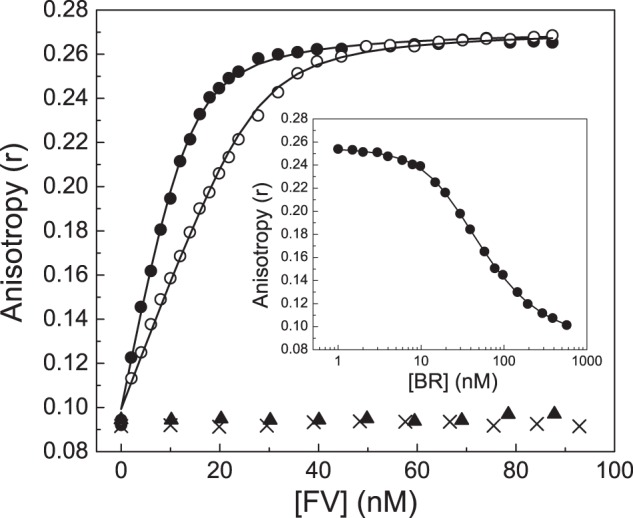
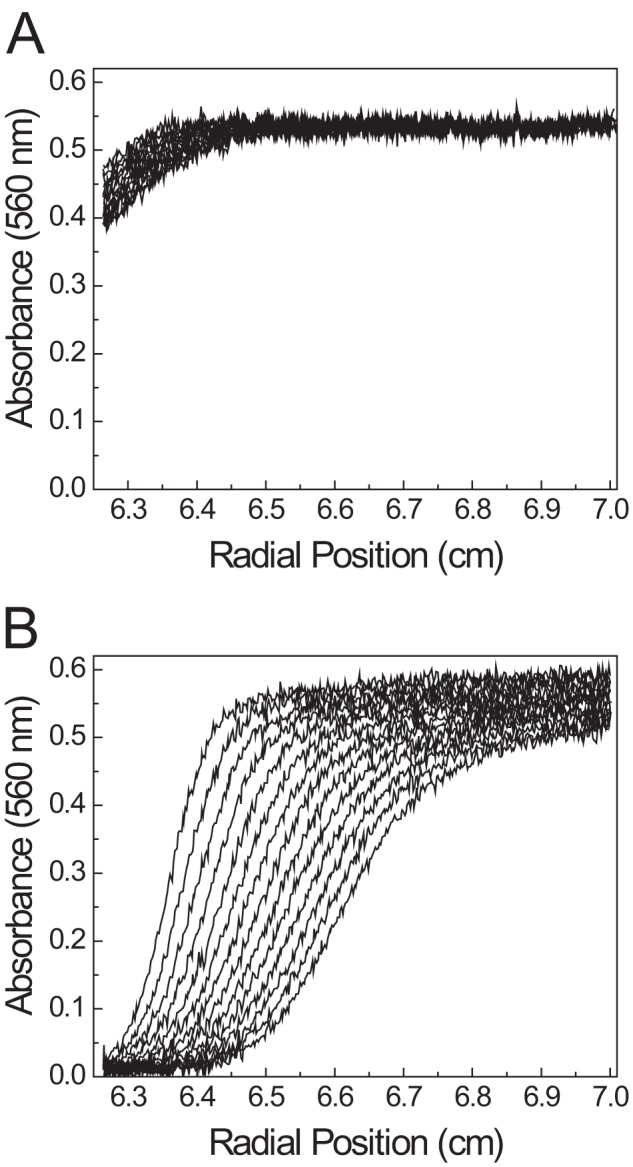
Coagulation factor V (FV) circulates as an inactive procofactor and is activated to FVa by proteolytic removal of a large inhibitory B-domain. Conserved basic and acidic sequences within the B-domain appear to play an important role in keeping FV as an inactive procofactor. Here, we utilized recombinant B-domain fragments to elucidate the mechanism of this FV autoinhibition. We show that a fragment encoding the basic region (BR) of the B-domain binds with high affinity to cofactor-like FV(a) variants that harbor an intact acidic region. Furthermore, the BR inhibits procoagulant function of the variants, thereby restoring the procofactor state. The BR competes with FXa for binding to FV(a), and limited proteolysis of the B-domain, specifically at Arg(1545), ablates BR binding to promote high affinity association between FVa and FXa. These results provide new insight into the mechanism by which the B-domain stabilizes FV as an inactive procofactor and reveal how limited proteolysis of FV progressively destabilizes key regulatory regions of the B-domain to produce an active form of the molecule.
Keywords: Autoinhibition; Coagulation Factors; Enzyme Mechanisms; Factor V; Factor Va; Hemostasis; Protein Engineering; Prothrombinase; Recombinant Protein Expression.
Figures
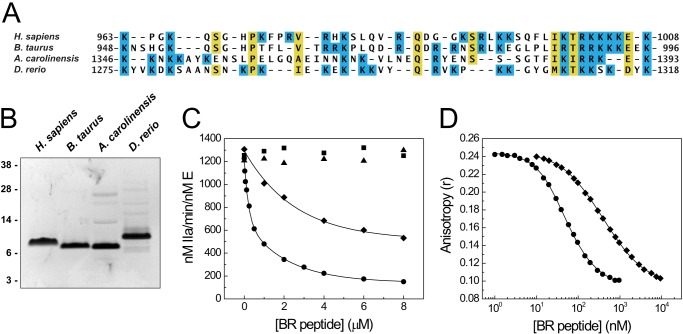
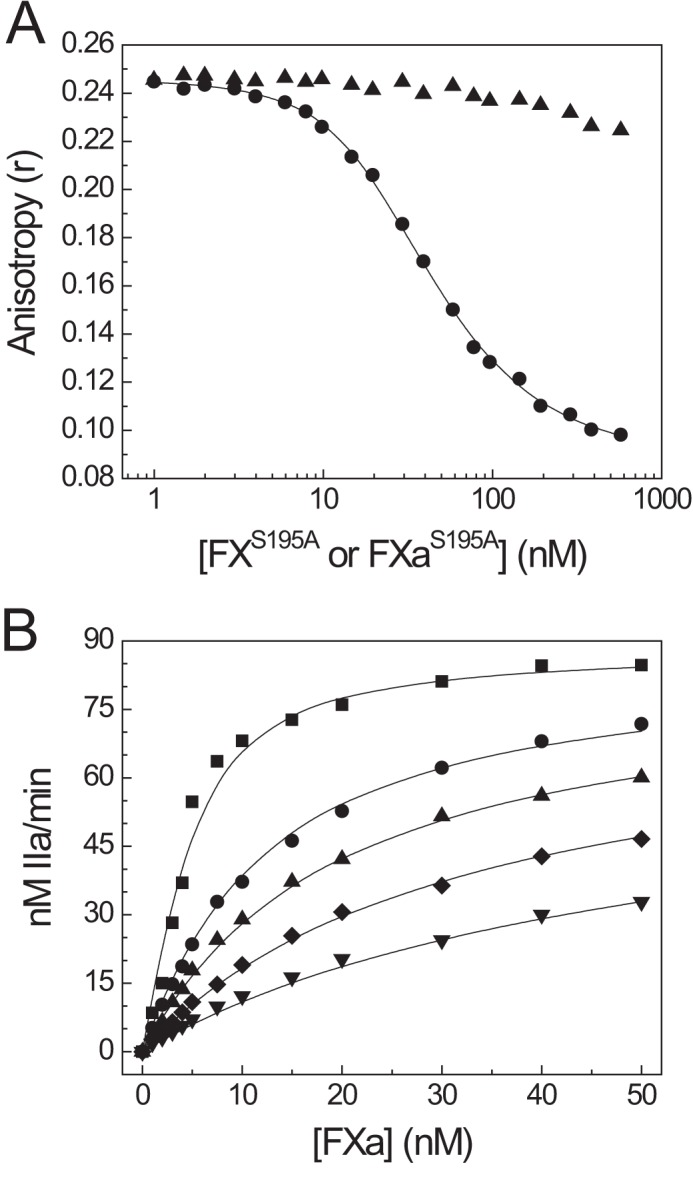
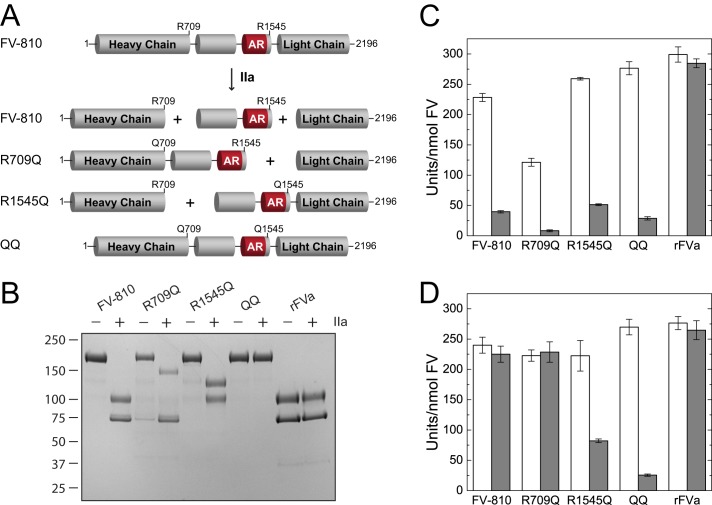
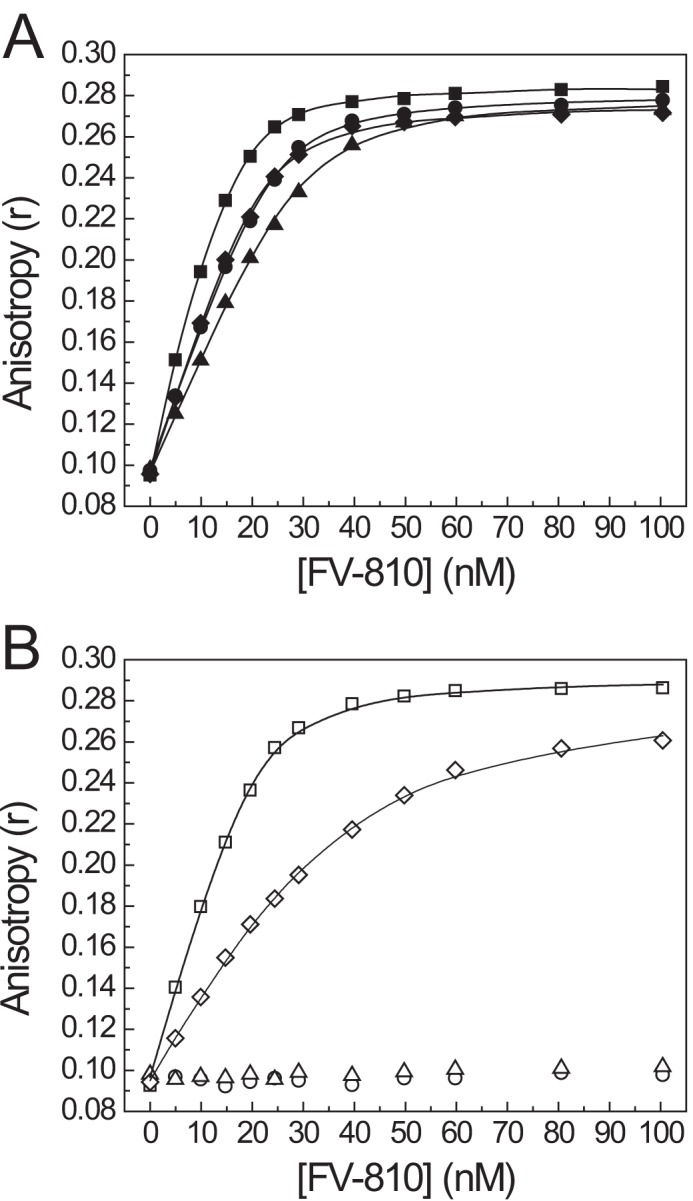
Similar articles
-
Regulation of factor V and factor V-short by TFPIα: Relationship between B-domain proteolysis and binding.J Biol Chem. 2021 Jan-Jun;296:100234. doi: 10.1074/jbc.RA120.016341. Epub 2021 Jan 7. J Biol Chem. 2021. PMID: 33376137 Free PMC article.
-
Factor Va-factor Xa interactions: molecular sites involved in enzyme:cofactor assembly.Scand J Clin Lab Invest Suppl. 2002;237:5-11. doi: 10.1080/003655102762377439. Scand J Clin Lab Invest Suppl. 2002. PMID: 12570161 Review.
-
Regulation of factor V by the anticoagulant protease activated protein C: Influence of the B-domain and TFPIα.J Biol Chem. 2022 Nov;298(11):102558. doi: 10.1016/j.jbc.2022.102558. Epub 2022 Sep 30. J Biol Chem. 2022. PMID: 36183835 Free PMC article.
-
Amino acid region 1000-1008 of factor V is a dynamic regulator for the emergence of procoagulant activity.J Biol Chem. 2013 Dec 27;288(52):37026-38. doi: 10.1074/jbc.M113.462374. Epub 2013 Oct 31. J Biol Chem. 2013. PMID: 24178294 Free PMC article.
-
The molecular basis of factor V and VIII procofactor activation.J Thromb Haemost. 2009 Dec;7(12):1951-61. doi: 10.1111/j.1538-7836.2009.03622.x. Epub 2009 Sep 18. J Thromb Haemost. 2009. PMID: 19765210 Free PMC article. Review.
Cited by
-
Biology of tissue factor pathway inhibitor.Blood. 2014 May 8;123(19):2934-43. doi: 10.1182/blood-2013-11-512764. Epub 2014 Mar 11. Blood. 2014. PMID: 24620349 Free PMC article. Review.
-
A hydrophobic patch (PLVIVG; 1481-1486) in the B-domain of factor V-short is crucial for its synergistic TFPIα-cofactor activity with protein S and for the formation of the FXa-inhibitory complex comprising FV-short, TFPIα, and protein S.J Thromb Haemost. 2022 May;20(5):1146-1157. doi: 10.1111/jth.15690. Epub 2022 Mar 16. J Thromb Haemost. 2022. PMID: 35247027 Free PMC article.
-
Blood coagulation factor Va's key interactive residues and regions for prothrombinase assembly and prothrombin binding.J Thromb Haemost. 2019 Aug;17(8):1229-1239. doi: 10.1111/jth.14487. Epub 2019 Jun 17. J Thromb Haemost. 2019. PMID: 31102425 Free PMC article. Review.
-
Cryo-EM structures of human coagulation factors V and Va.Blood. 2021 Jun 3;137(22):3137-3144. doi: 10.1182/blood.2021010684. Blood. 2021. PMID: 33684942 Free PMC article.
-
Factor V-short and protein S as synergistic tissue factor pathway inhibitor (TFPIα) cofactors.Res Pract Thromb Haemost. 2017 Dec 20;2(1):114-124. doi: 10.1002/rth2.12057. eCollection 2018 Jan. Res Pract Thromb Haemost. 2017. PMID: 30046712 Free PMC article.
References
-
- Mann K. G., Nesheim M. E., Church W. R., Haley P., Krishnaswamy S. (1990) Surface-dependent reactions of the vitamin K-dependent enzyme complexes. Blood 76, 1–16 - PubMed
-
- Mann K. G., Kalafatis M. (2003) Factor V: a combination of Dr. Jekyll and Mr. Hyde. Blood 101, 20–30 - PubMed
-
- Esmon C. T., Owen W. G., Duiguid D. L., Jackson C. M. (1973) The action of thrombin on blood clotting factor V: conversion of factor V to a prothrombin-binding protein. Biochim. Biophys. Acta 310, 289–294 - PubMed
-
- Foster W. B., Nesheim M. E., Mann K. G. (1983) The Factor Xa-catalyzed activation of Factor V. J. Biol. Chem. 258, 13970–13977 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
