

DNA methylation presents distinct binding sites for human transcription factors
- PMID: 24015356
- PMCID: PMC3762332
- DOI: 10.7554/eLife.00726
DNA methylation presents distinct binding sites for human transcription factors
Abstract
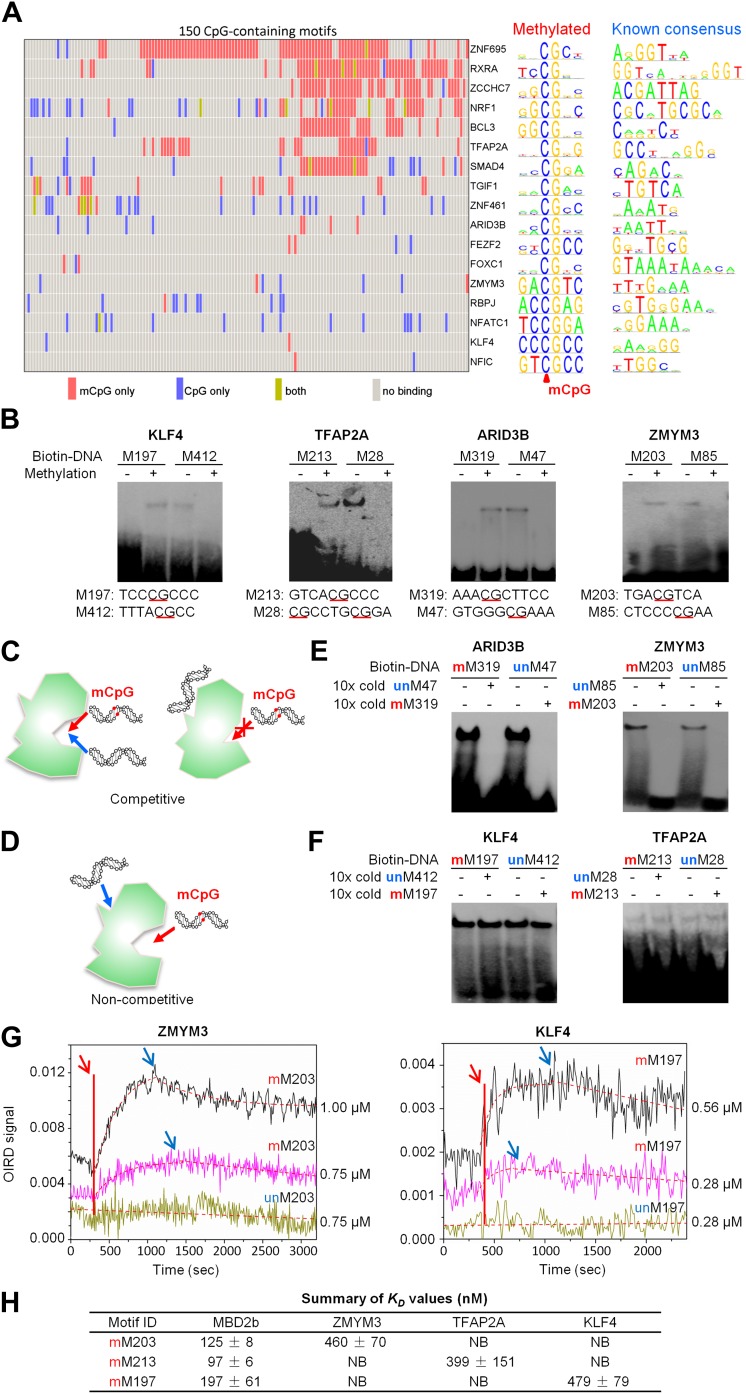
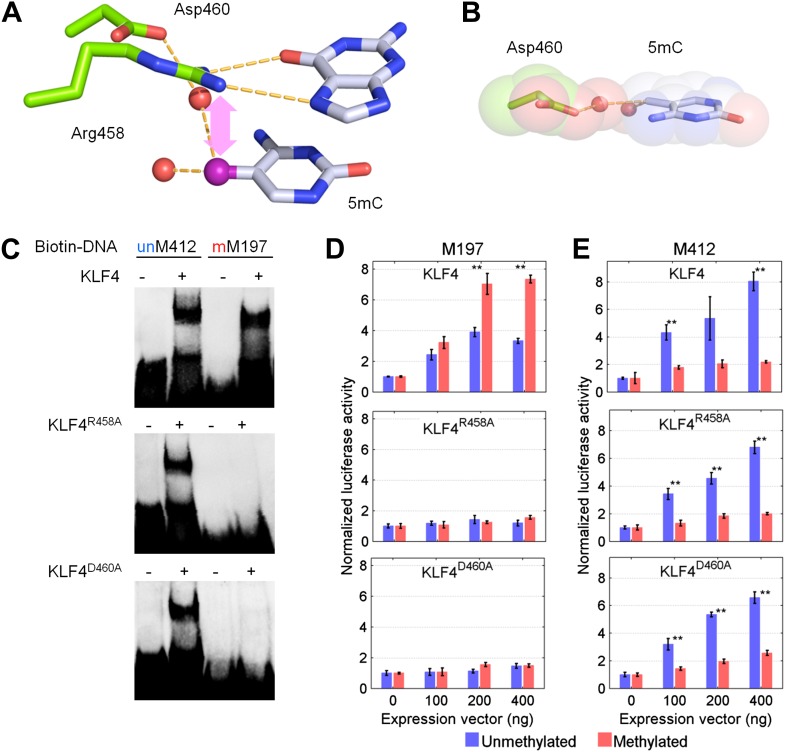
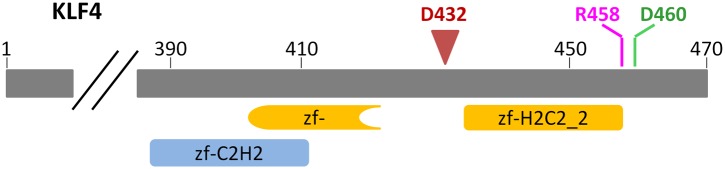
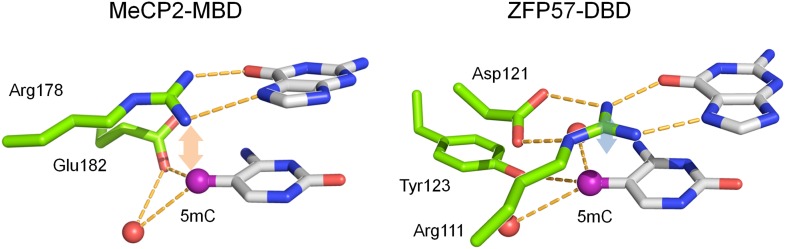
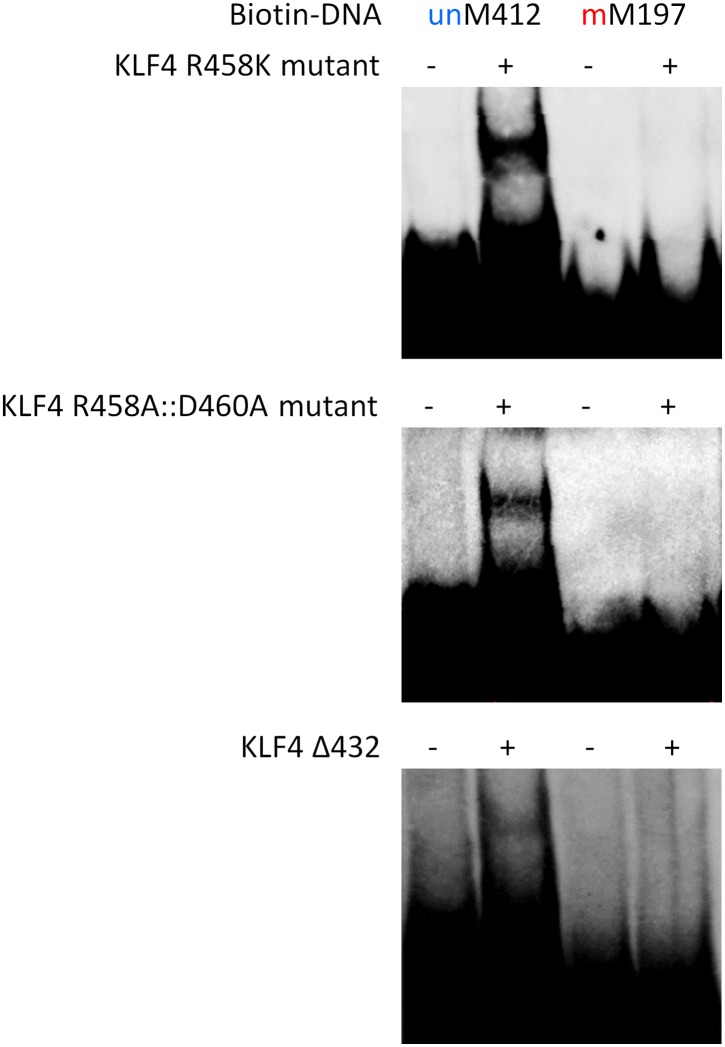
DNA methylation, especially CpG methylation at promoter regions, has been generally considered as a potent epigenetic modification that prohibits transcription factor (TF) recruitment, resulting in transcription suppression. Here, we used a protein microarray-based approach to systematically survey the entire human TF family and found numerous purified TFs with methylated CpG (mCpG)-dependent DNA-binding activities. Interestingly, some TFs exhibit specific binding activity to methylated and unmethylated DNA motifs of distinct sequences. To elucidate the underlying mechanism, we focused on Kruppel-like factor 4 (KLF4), and decoupled its mCpG- and CpG-binding activities via site-directed mutagenesis. Furthermore, KLF4 binds specific methylated or unmethylated motifs in human embryonic stem cells in vivo. Our study suggests that mCpG-dependent TF binding activity is a widespread phenomenon and provides a new framework to understand the role and mechanism of TFs in epigenetic regulation of gene transcription. DOI:http://dx.doi.org/10.7554/eLife.00726.001.
Keywords: DNA methylation; Human; epigenetics; protein microarray; protein-DNA interactions; transcription factors; transcription regulation.
Conflict of interest statement
The authors declare that no competing interests exist.
Figures


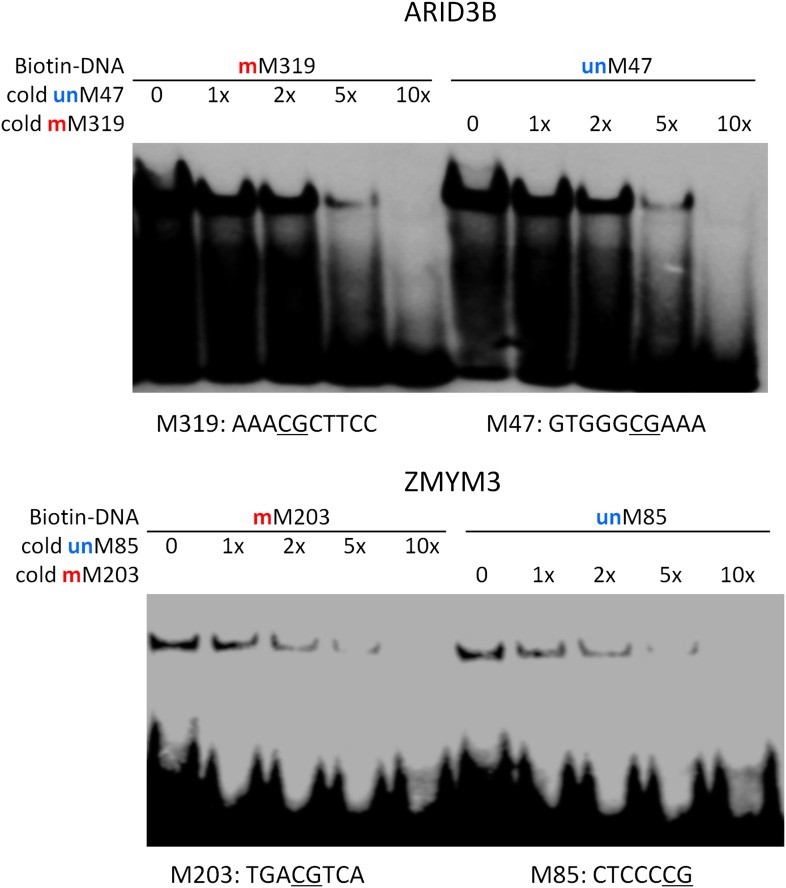
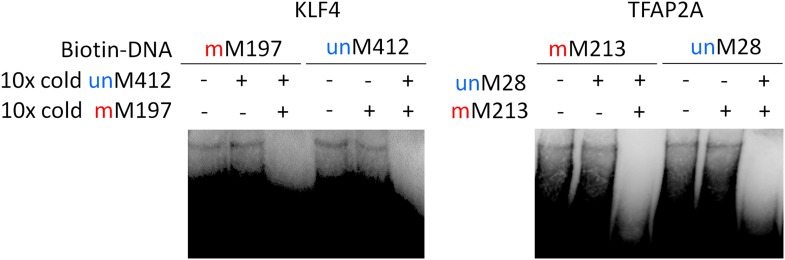
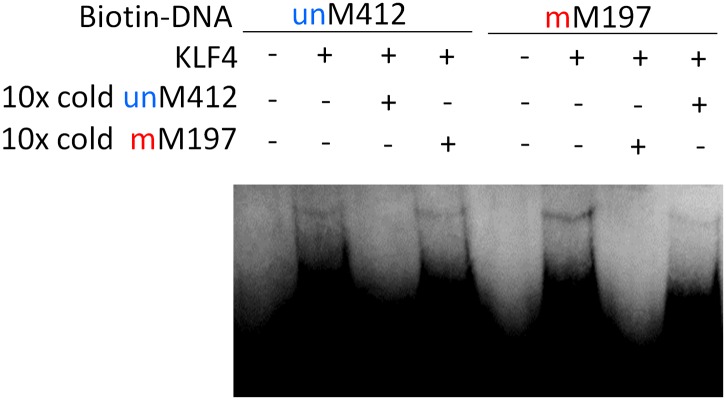
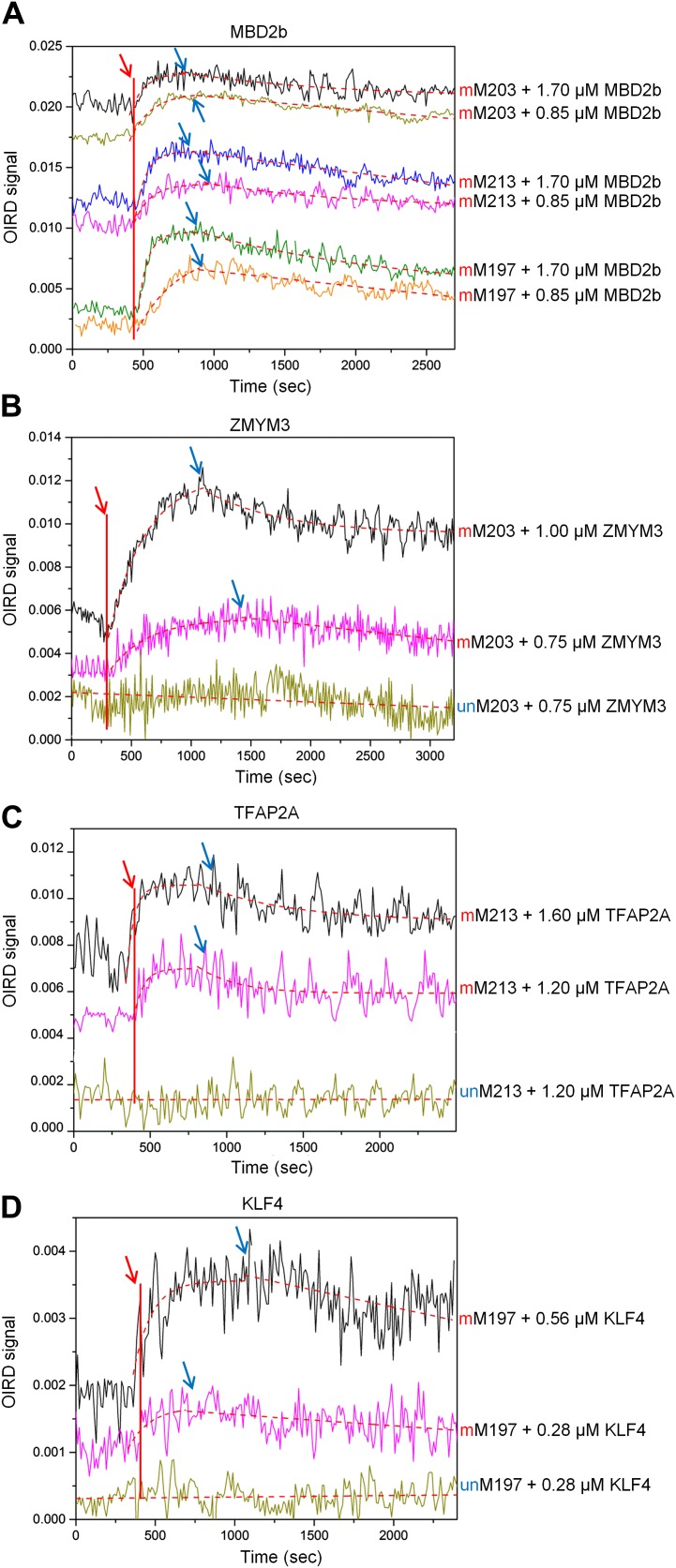

Comment in
-
Making the most of methylation.Elife. 2013 Sep 3;2:e01387. doi: 10.7554/eLife.01387. Elife. 2013. PMID: 24015362 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
