

Mechanism of synergistic activation of Arp2/3 complex by cortactin and N-WASP
- PMID: 24015358
- PMCID: PMC3762189
- DOI: 10.7554/eLife.00884
Mechanism of synergistic activation of Arp2/3 complex by cortactin and N-WASP
Abstract
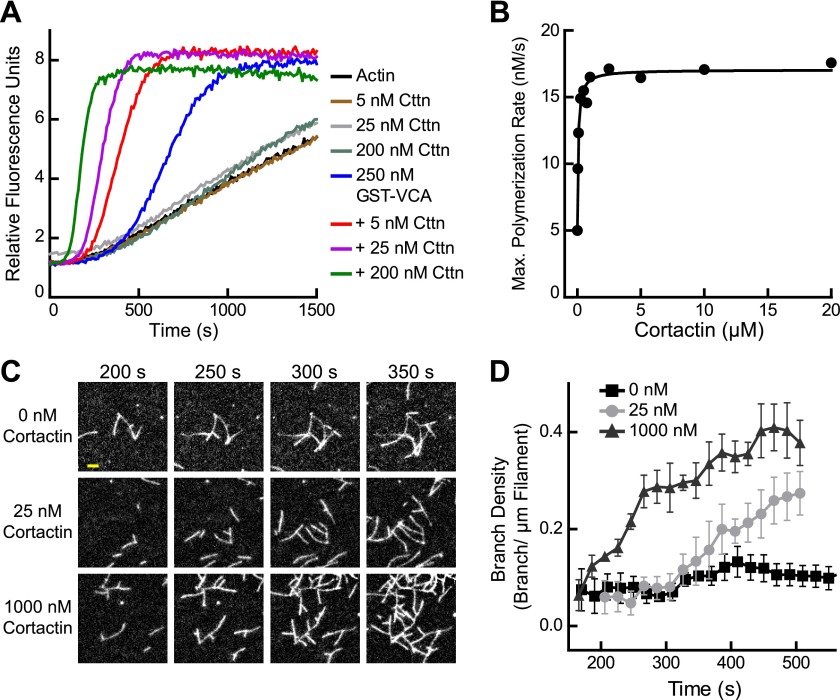
Nucleation promoting factors (NPFs) initiate branched actin network assembly by activating Arp2/3 complex, a branched actin filament nucleator. Cellular actin networks contain multiple NPFs, but how they coordinately regulate Arp2/3 complex is unclear. Cortactin is an NPF that activates Arp2/3 complex weakly on its own, but with WASP/N-WASP, another class of NPFs, potently activates. We dissect the mechanism of synergy and propose a model in which cortactin displaces N-WASP from nascent branches as a prerequisite for nucleation. Single-molecule imaging revealed that unlike WASP/N-WASP, cortactin remains bound to junctions during nucleation, and specifically targets junctions with a ∼160-fold increased on rate over filament sides. N-WASP must be dimerized for potent synergy, and targeted mutations indicate release of dimeric N-WASP from nascent branches limits nucleation. Mathematical modeling shows cortactin-mediated displacement but not N-WASP recycling or filament recruitment models can explain synergy. Our results provide a molecular basis for coordinate Arp2/3 complex regulation. DOI:http://dx.doi.org/10.7554/eLife.00884.001.
Keywords: Arp2/3; Mouse; N-WASP; WASP; actin; cortactin; single molecule.
Conflict of interest statement
The authors declare that no competing interests exist.
Figures












References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
