

Proteomic and virus-induced gene silencing (VIGS) Analyses reveal that gossypol, brassinosteroids, and jasmonic acid contribute to the resistance of cotton to Verticillium dahliae
- PMID: 24019146
- PMCID: PMC3861717
- DOI: 10.1074/mcp.M113.031013
Proteomic and virus-induced gene silencing (VIGS) Analyses reveal that gossypol, brassinosteroids, and jasmonic acid contribute to the resistance of cotton to Verticillium dahliae
Abstract
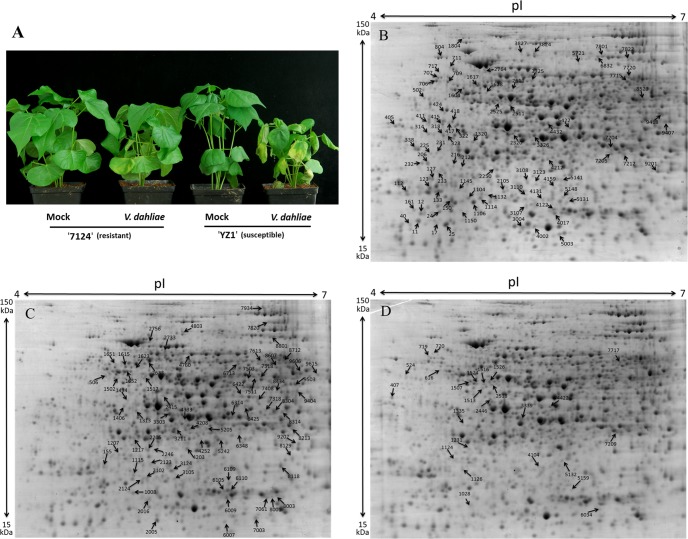
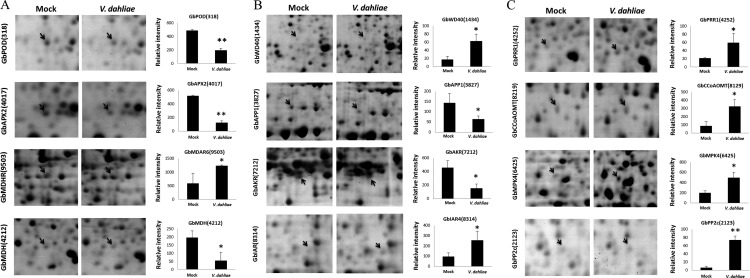
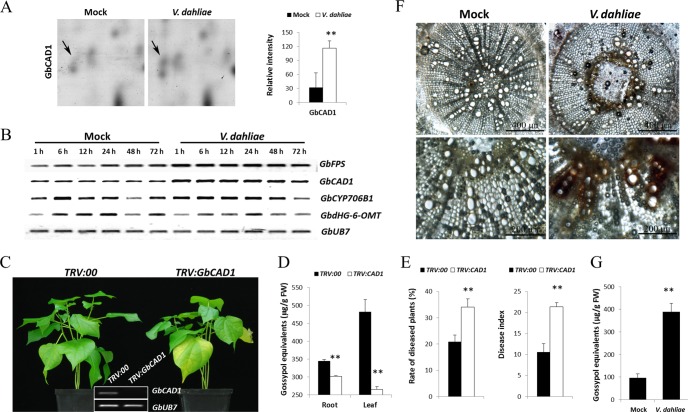
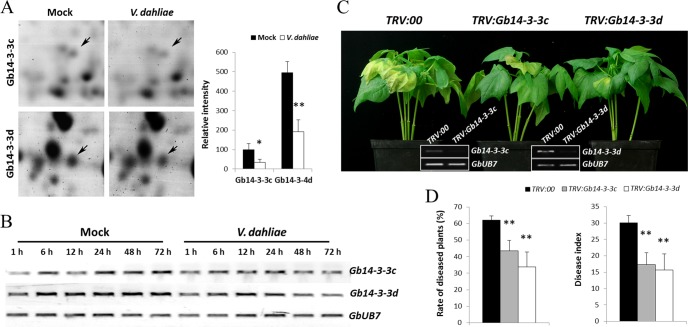
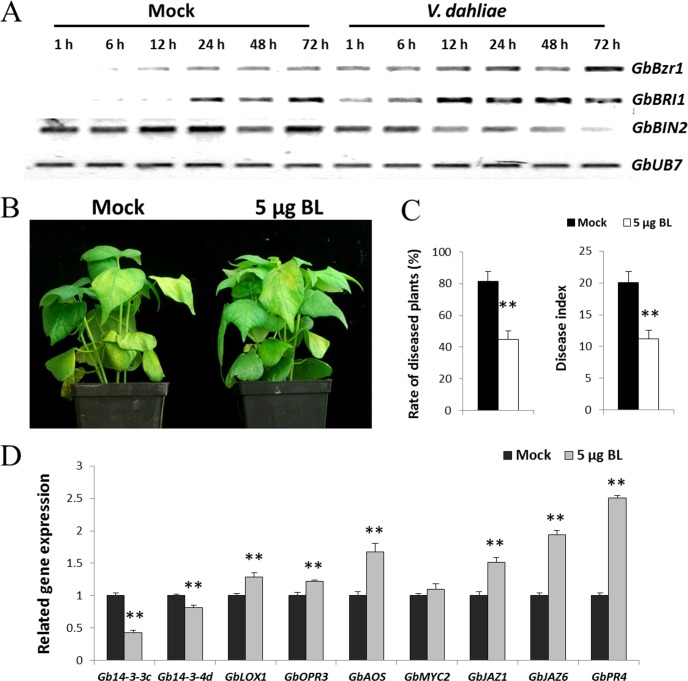
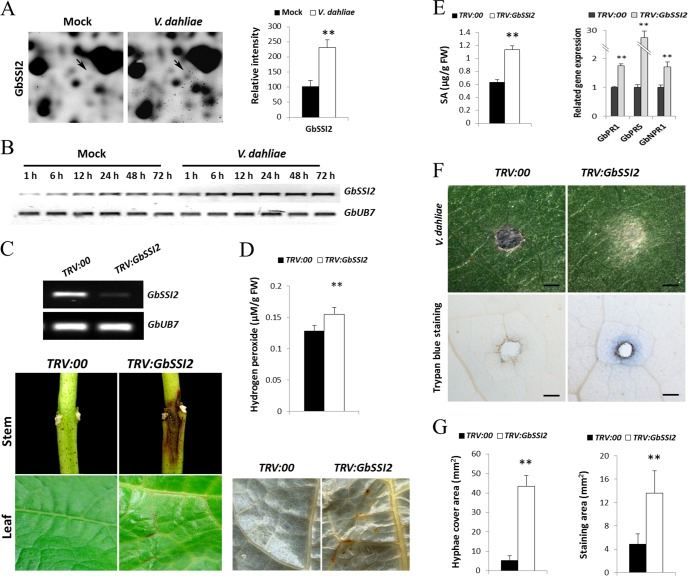
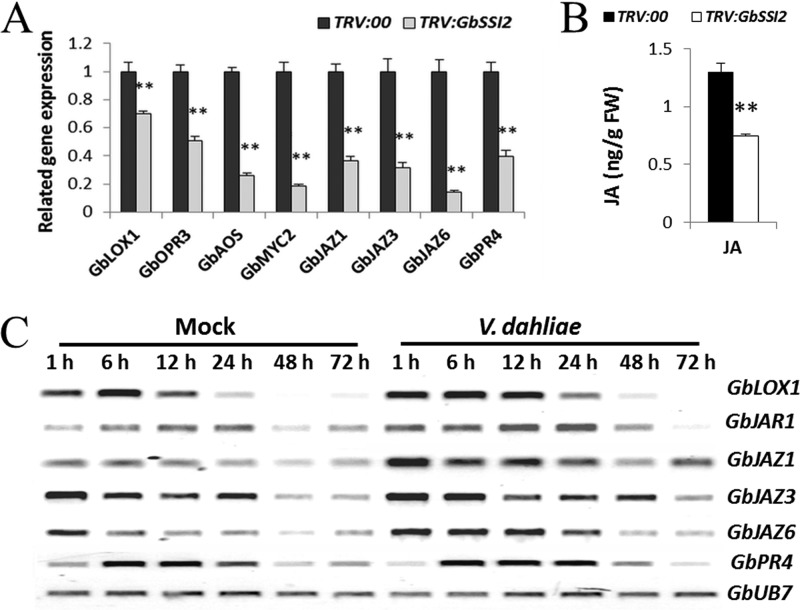
Verticillium wilt causes massive annual losses of cotton yield, but the mechanism of cotton resistance to Verticillium dahliae is complex and poorly understood. In this study, a comparative proteomic analysis was performed in resistant cotton (Gossypium barbadense cv7124) on infection with V. dahliae. A total of 188 differentially expressed proteins were identified by mass spectrometry (MALDI-TOF/TOF) analysis and could be classified into 17 biological processes based on Gene Ontology annotation. Most of these proteins were implicated in stimulus response, cellular processes and metabolic processes. Based on the proteomic analysis, several genes involved in secondary metabolism, reactive oxygen burst and phytohormone signaling pathways were identified for further physiological and molecular analysis. The roles of the corresponding genes were further characterized by employing virus-induced gene silencing (VIGS). Based on the results, we suggest that the production of gossypol is sufficient to affect the cotton resistance to V. dahliae. Silencing of GbCAD1, a key enzyme involving in gossypol biosynthesis, compromised cotton resistance to V. dahliae. Reactive oxygen species and salicylic acid signaling may be also implicated as regulators in cotton responsive to V. dahliae according to the analysis of GbSSI2, an important regulator in the crosstalk between salicylic acid and jasmonic acid signal pathways. Moreover, brassinosteroids and jasmonic acid signaling may play essential roles in the cotton disease resistance to V. dahliae. The brassinosteroids signaling was activated in cotton on inoculation with V. dahliae and the disease resistance of cotton was enhanced after exogenous application of brassinolide. Meanwhile, jasmonic acid signaling was also activated in cotton after inoculation with V. dahliae and brassinolide application. These data provide highlights in the molecular basis of cotton resistance to V. dahliae.
Figures
Similar articles
-
Suppression of the homeobox gene HDTF1 enhances resistance to Verticillium dahliae and Botrytis cinerea in cotton.J Integr Plant Biol. 2016 May;58(5):503-13. doi: 10.1111/jipb.12432. Epub 2015 Nov 21. J Integr Plant Biol. 2016. PMID: 26407676
-
Functional characterization of cotton genes responsive to Verticillium dahliae through bioinformatics and reverse genetics strategies.J Exp Bot. 2014 Dec;65(22):6679-92. doi: 10.1093/jxb/eru393. Epub 2014 Oct 17. J Exp Bot. 2014. PMID: 25326626 Free PMC article.
-
Melatonin enhances cotton immunity to Verticillium wilt via manipulating lignin and gossypol biosynthesis.Plant J. 2019 Nov;100(4):784-800. doi: 10.1111/tpj.14477. Epub 2019 Aug 31. Plant J. 2019. PMID: 31349367 Free PMC article.
-
Insights to Gossypium defense response against Verticillium dahliae: the Cotton Cancer.Funct Integr Genomics. 2023 May 1;23(2):142. doi: 10.1007/s10142-023-01065-5. Funct Integr Genomics. 2023. PMID: 37121989 Review.
-
An Overview of the Molecular Genetics of Plant Resistance to the Verticillium Wilt Pathogen Verticillium dahliae.Int J Mol Sci. 2020 Feb 7;21(3):1120. doi: 10.3390/ijms21031120. Int J Mol Sci. 2020. PMID: 32046212 Free PMC article. Review.
Cited by
-
Development of an efficient and precise adenine base editor (ABE) with expanded target range in allotetraploid cotton (Gossypium hirsutum).BMC Biol. 2022 Feb 15;20(1):45. doi: 10.1186/s12915-022-01232-3. BMC Biol. 2022. PMID: 35164736 Free PMC article.
-
GhCPK33 Negatively Regulates Defense against Verticillium dahliae by Phosphorylating GhOPR3.Plant Physiol. 2018 Oct;178(2):876-889. doi: 10.1104/pp.18.00737. Epub 2018 Aug 27. Plant Physiol. 2018. PMID: 30150302 Free PMC article.
-
The Calcium Sensor CBL2 and Its Interacting Kinase CIPK6 Are Involved in Plant Sugar Homeostasis via Interacting with Tonoplast Sugar Transporter TST2.Plant Physiol. 2020 May;183(1):236-249. doi: 10.1104/pp.19.01368. Epub 2020 Mar 5. Plant Physiol. 2020. PMID: 32139477 Free PMC article.
-
Potential of Endophytic Fungi Isolated from Cotton Roots for Biological Control against Verticillium Wilt Disease.PLoS One. 2017 Jan 20;12(1):e0170557. doi: 10.1371/journal.pone.0170557. eCollection 2017. PLoS One. 2017. PMID: 28107448 Free PMC article.
-
Hormone Signaling and Its Interplay With Development and Defense Responses in Verticillium-Plant Interactions.Front Plant Sci. 2020 Nov 4;11:584997. doi: 10.3389/fpls.2020.584997. eCollection 2020. Front Plant Sci. 2020. PMID: 33250913 Free PMC article. Review.
References
-
- Sal'kova E. G., Guseva N. N. (1965) The role of pectolytic enzymes of the verticillium dahliae fungus in the development of cotton wilt. Dokl. Akad. Nauk SSSR 163, 515–522 - PubMed
-
- Cai Y. F., He X. H., Mo J. C., Sun Q., Yang J. P., Liu J. G. (2009) Molecular research and genetic engineering of resistance to Verticillium wilt in cotton: A review. Afr. J. Biotechnol. 8, 7363–7372
-
- Zhang J., Sanogo S., Flynn R., Baral J., Bajaj S., Hughs S. E., Percy R. (2012) Germplasm evaluation and transfer of Verticillium wilt resistance from Pima (Gossypium barbadense) to Upland cotton (G. hirsutum). Euphytica 187, 147–160
-
- Aguado A., Santos B. D. L., Blanco C., Romero F. (2008) Study of gene effects for cotton yield and Verticillium wilt tolerance in cotton plant (Gossypium hirsutum L.). Field Crops Res. 107, 78–86
-
- Jiang F., Zhao J., Zhou L., Guo W., Zhang T. (2009) Molecular mapping of Verticillium wilt resistance QTL clustered on chromosomes D7 and D9 in upland cotton. Sci. China C Life Sci. 52, 872–884 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
