

Coalescence and genetic diversity in sexual populations under selection
- PMID: 24019480
- PMCID: PMC3785765
- DOI: 10.1073/pnas.1309697110
Coalescence and genetic diversity in sexual populations under selection
Abstract
In sexual populations, selection operates neither on the whole genome, which is repeatedly taken apart and reassembled by recombination, nor on individual alleles that are tightly linked to the chromosomal neighborhood. The resulting interference between linked alleles reduces the efficiency of selection and distorts patterns of genetic diversity. Inference of evolutionary history from diversity shaped by linked selection requires an understanding of these patterns. Here, we present a simple but powerful scaling analysis identifying the unit of selection as the genomic "linkage block" with a characteristic length, , determined in a self-consistent manner by the condition that the rate of recombination within the block is comparable to the fitness differences between different alleles of the block. We find that an asexual model with the strength of selection tuned to that of the linkage block provides an excellent description of genetic diversity and the site frequency spectra compared with computer simulations. This linkage block approximation is accurate for the entire spectrum of strength of selection and is particularly powerful in scenarios with many weakly selected loci. The latter limit allows us to characterize coalescence, genetic diversity, and the speed of adaptation in the infinitesimal model of quantitative genetics.
Keywords: Bolthausen–Sznitman coalescent; Hill–Robertson interference; genealogy.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
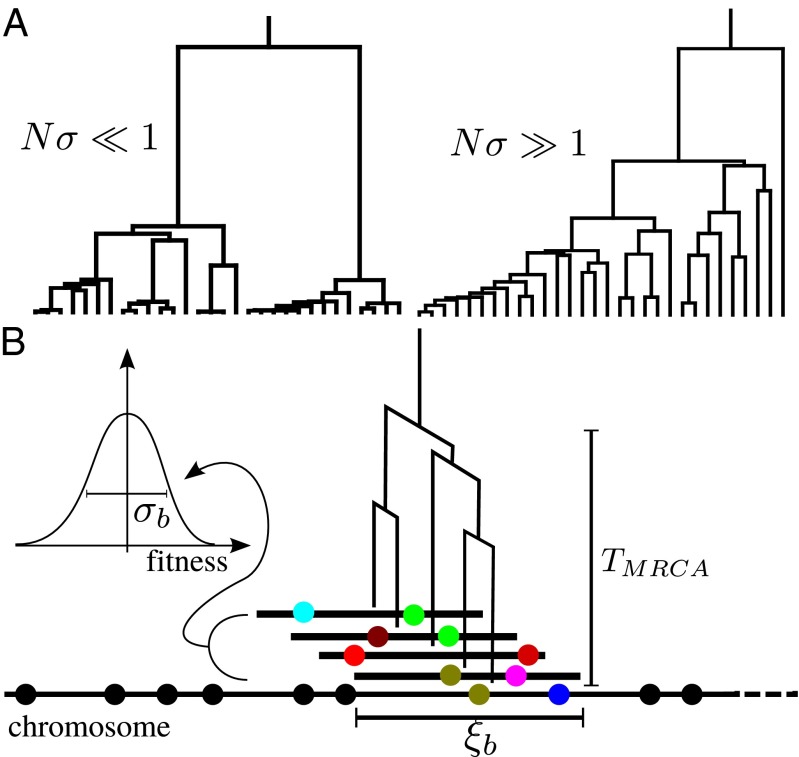
 share most of their genealogical history, (i.e., have trees similar to the one indicated in the center of the segment where TMRCA is the time to the most recent common ancestor). Coalescence within this segment of length
share most of their genealogical history, (i.e., have trees similar to the one indicated in the center of the segment where TMRCA is the time to the most recent common ancestor). Coalescence within this segment of length  is either neutral or driven by the fitness differences between different haplotypes spanning these segments. (Inset) Fitness distribution of these haplotype blocks is indicated. Distant parts of the chromosome are in linkage equilibrium, and the tree changes as one moves along the chromosome. The succession of changing trees is the ancestral recombination graph.
is either neutral or driven by the fitness differences between different haplotypes spanning these segments. (Inset) Fitness distribution of these haplotype blocks is indicated. Distant parts of the chromosome are in linkage equilibrium, and the tree changes as one moves along the chromosome. The succession of changing trees is the ancestral recombination graph.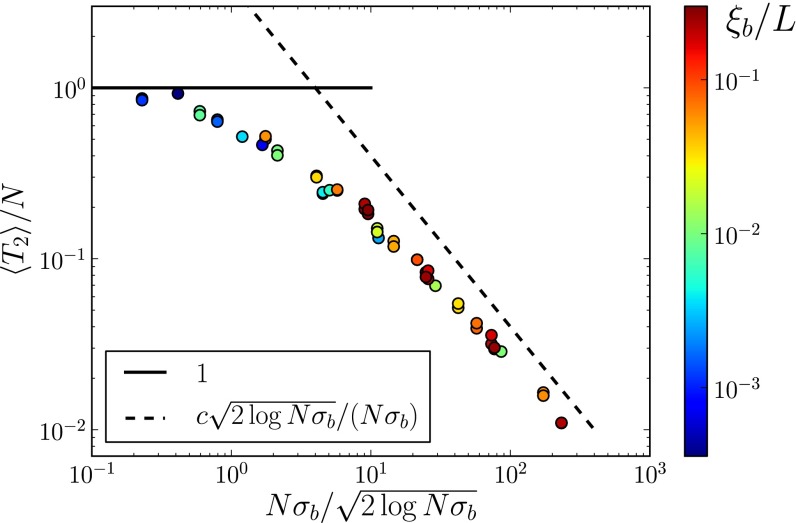
 relative to the neutral expectation as a function of
relative to the neutral expectation as a function of  determined using Eq. 5. For
determined using Eq. 5. For  ,
,  , whereas
, whereas  otherwise.
otherwise.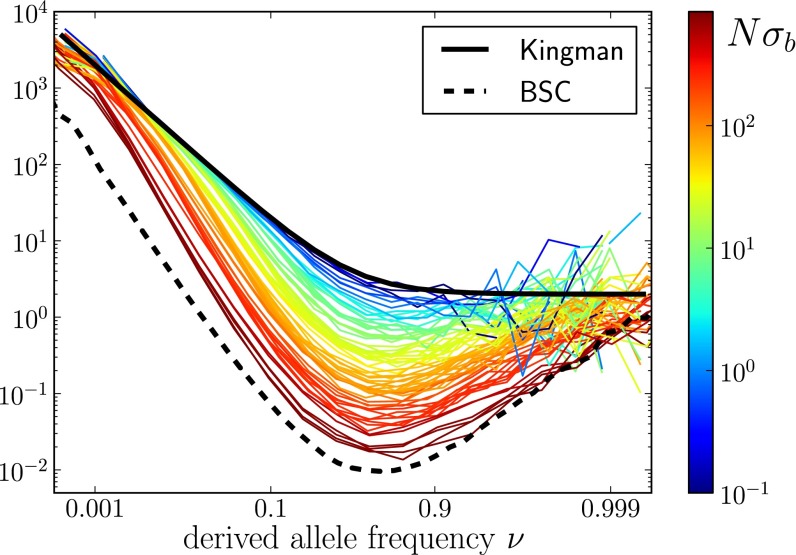
 , for a large number of parameter combinations. Color indicates the value of
, for a large number of parameter combinations. Color indicates the value of  . For large
. For large  , the SFSs display the nonmonotonicity characteristic of the BSC (dashed line), whereas the SFSs are described well by the prediction from Kingman’s coalescent (solid line) if
, the SFSs display the nonmonotonicity characteristic of the BSC (dashed line), whereas the SFSs are described well by the prediction from Kingman’s coalescent (solid line) if  . The BSC curve serves as a guide to the eye because its normalization depends on
. The BSC curve serves as a guide to the eye because its normalization depends on  .
.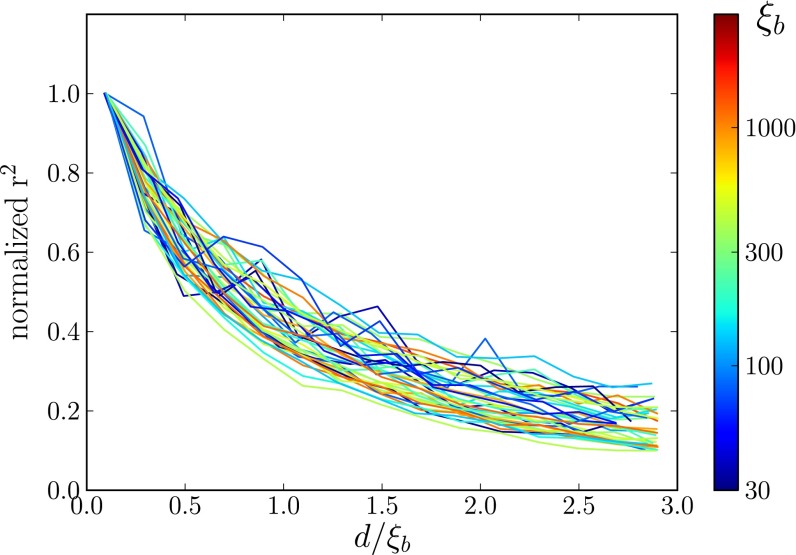
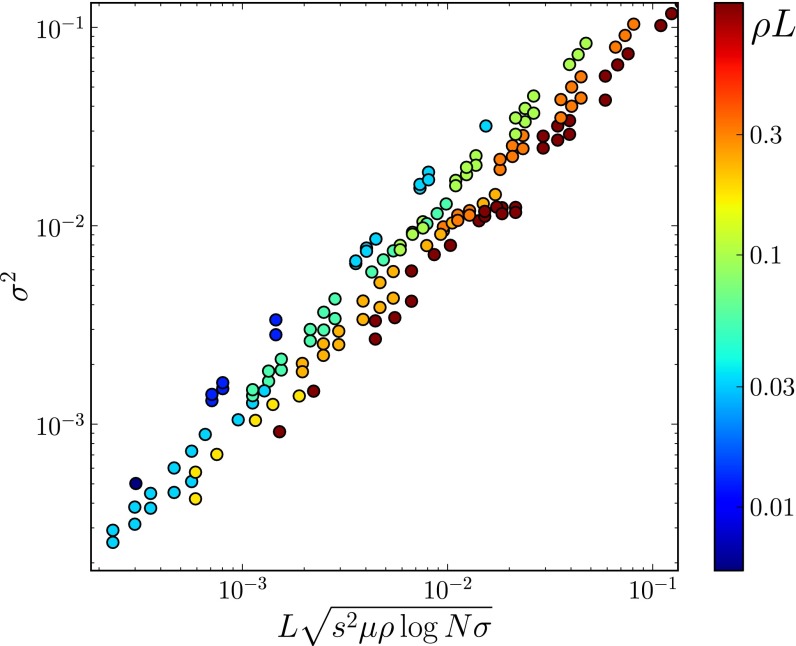
 , between pairs of loci at different distances (the curves are normalized to their value at zero distance). The x axis shows the distance between loci d rescaled by
, between pairs of loci at different distances (the curves are normalized to their value at zero distance). The x axis shows the distance between loci d rescaled by  determined using Eq. 2, with t equal to the measured pair coalescence time. After this rescaling, the distance dependence of all simulations follows approximately the same master curve, which shows that LD extends for
determined using Eq. 2, with t equal to the measured pair coalescence time. After this rescaling, the distance dependence of all simulations follows approximately the same master curve, which shows that LD extends for  .
.
References
-
- Gerrish PJ, Lenski RE. The fate of competing beneficial mutations in an asexual population. Genetica. 1998;102-103(1-6):127–144. - PubMed
-
- Neher RA. Genetic draft, selective interference, and population genetics of rapid adaptation. Annu Rev Ecol Evol Syst. 2013:44. in press.
-
- Hill WG, Robertson A. The effect of linkage on limits to artificial selection. Genet Res. 1966;8(3):269–294. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
