

Dissection of Cdk1-cyclin complexes in vivo
- PMID: 24019491
- PMCID: PMC3785786
- DOI: 10.1073/pnas.1305420110
Dissection of Cdk1-cyclin complexes in vivo
Abstract
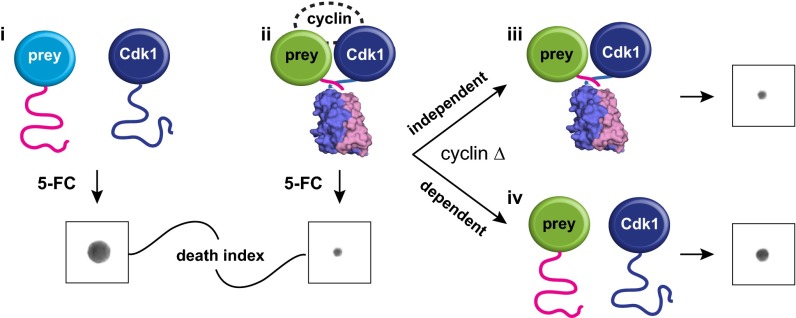
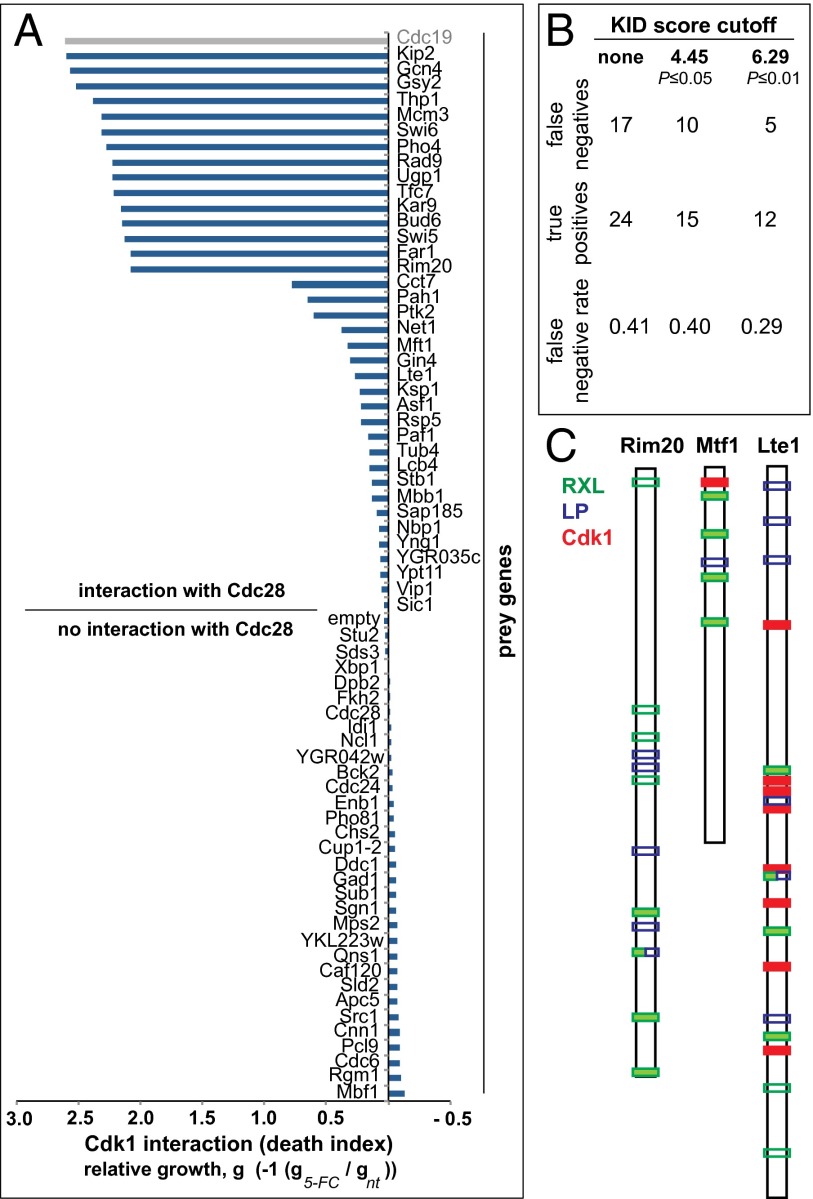
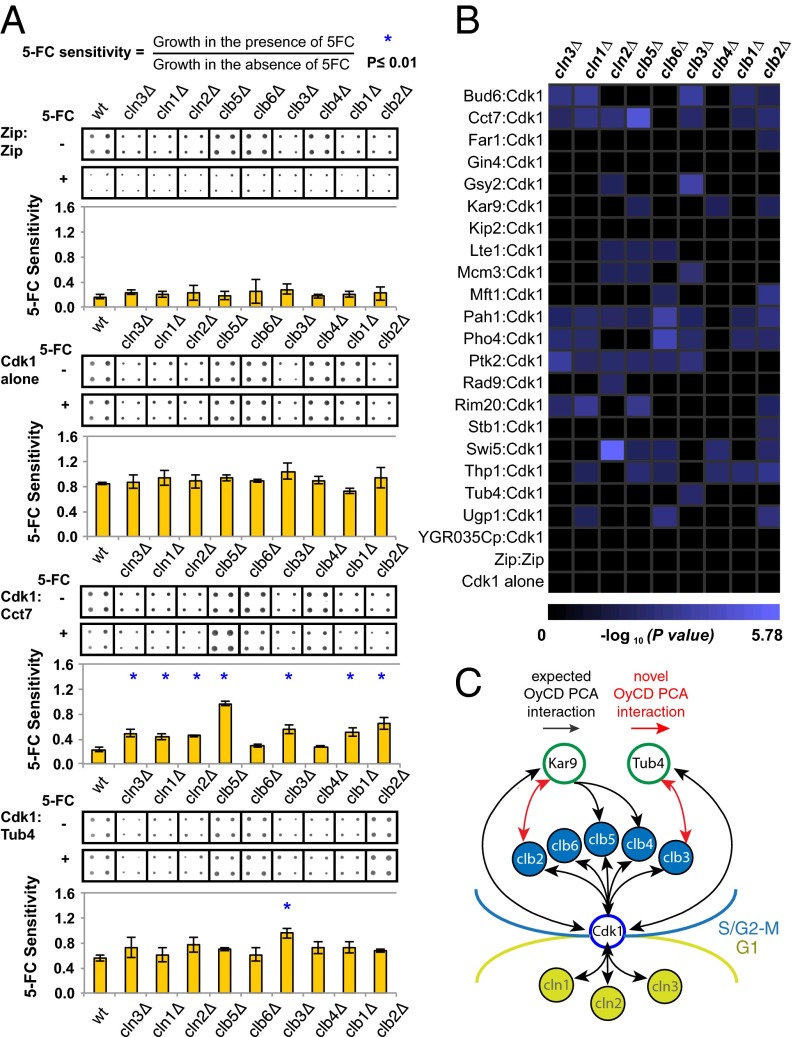
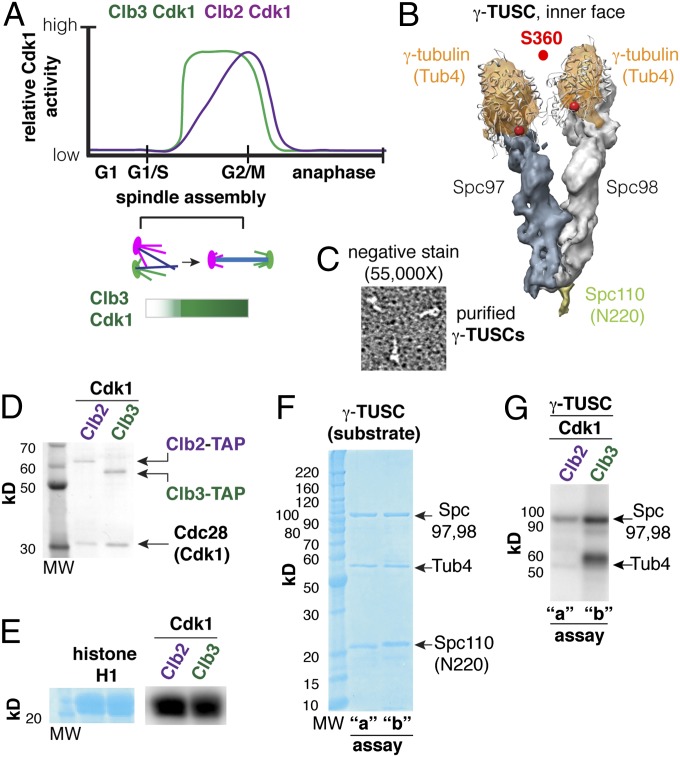
Cyclin-dependent kinases (Cdks) are regulatory enzymes with temporal and spatial selectivity for their protein substrates that are governed by cell cycle-regulated cyclin subunits. Specific cyclin-Cdk complexes bind to and phosphorylate target proteins, coupling their activity to cell cycle states. The identification of specific cyclin-Cdk substrates is challenging and so far, has largely been achieved through indirect correlation or use of in vitro techniques. Here, we use a protein-fragment complementation assay based on the optimized yeast cytosine deaminase to systematically identify candidate substrates of budding yeast Saccharomyces cerevisiae Cdk1 and show dependency on one or more regulatory cyclins. We identified known and candidate cyclin dependencies for many predicted protein kinase Cdk1 targets and showed elusory Clb3-Cdk1-specific phosphorylation of γ-tubulin, thus establishing the timing of this event in controlling assembly of the mitotic spindle. Our strategy can be generally applied to identify substrates and accessory subunits of multisubunit protein complexes.
Keywords: cyclin specificity; in vivo enzyme complexes screen.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Thornton BR, Toczyski DP. Precise destruction: An emerging picture of the APC. Genes Dev. 2006;20(22):3069–3078. - PubMed
-
- Cohen P. The structure and regulation of protein phosphatases. Annu Rev Biochem. 1989;58(1989):453–508. - PubMed
-
- Morgan DO. Cyclin-dependent kinases: Engines, clocks, and microprocessors. Annu Rev Cell Dev Biol. 1997;13(1997):261–291. - PubMed
-
- Janssens V, Longin S, Goris J. PP2A holoenzyme assembly: In cauda venenum (the sting is in the tail) Trends Biochem Sci. 2008;33(3):113–121. - PubMed
-
- Morgan DO. Principles of CDK regulation. Nature. 1995;374(6518):131–134. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
