

Lipid absorption defects in intestine-specific microsomal triglyceride transfer protein and ATP-binding cassette transporter A1-deficient mice
- PMID: 24019513
- PMCID: PMC3798507
- DOI: 10.1074/jbc.M113.501247
Lipid absorption defects in intestine-specific microsomal triglyceride transfer protein and ATP-binding cassette transporter A1-deficient mice
Abstract
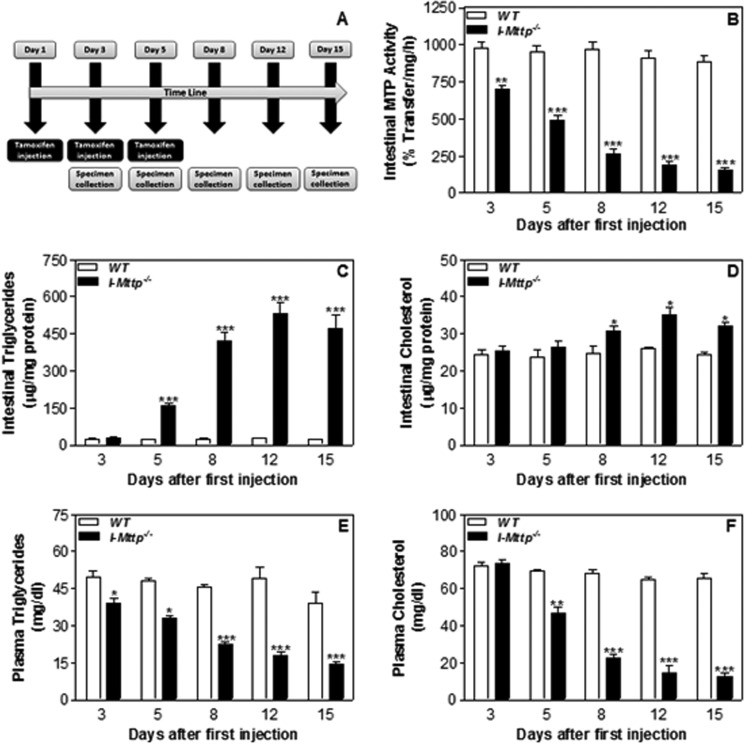
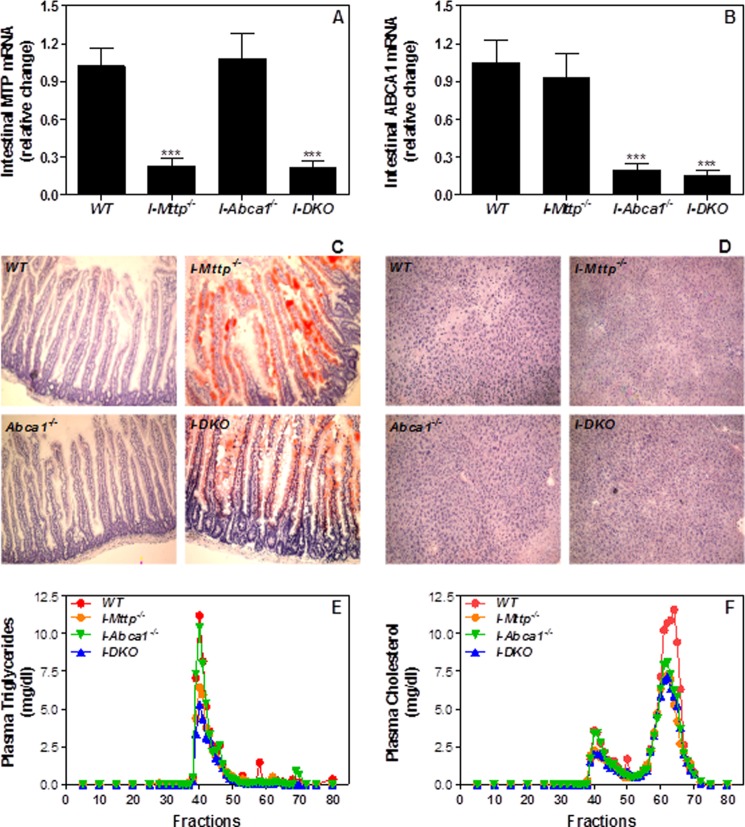
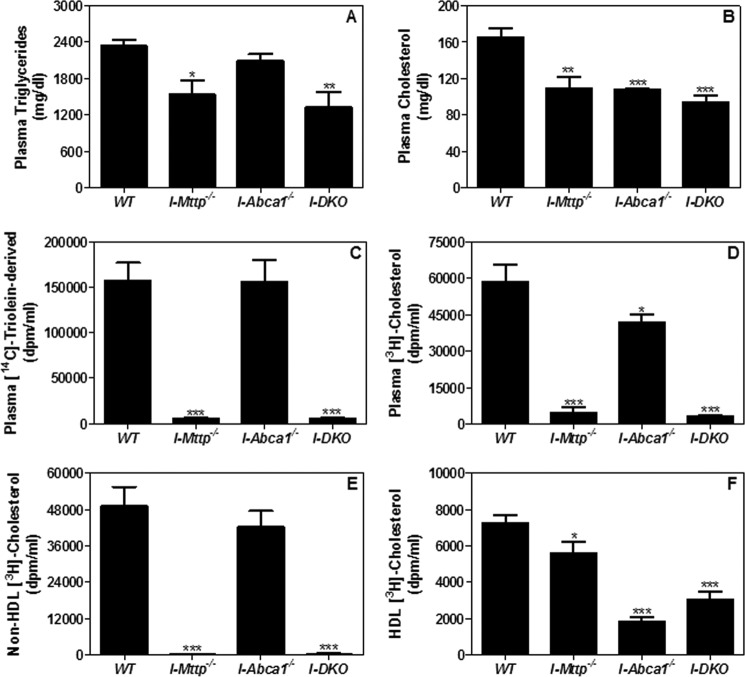
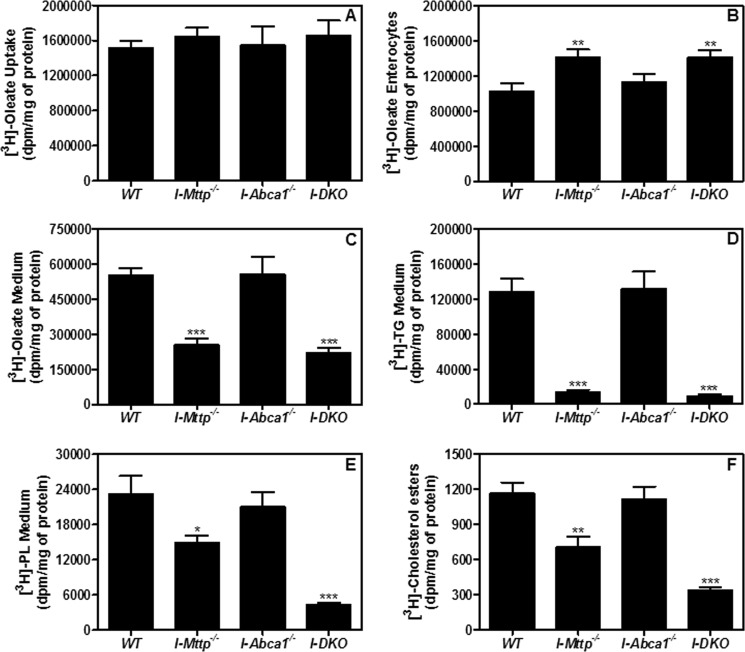
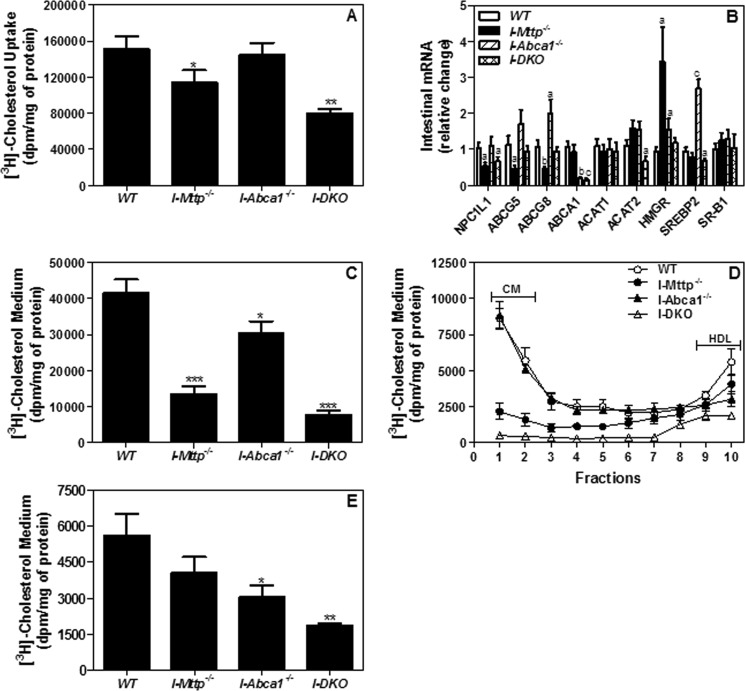
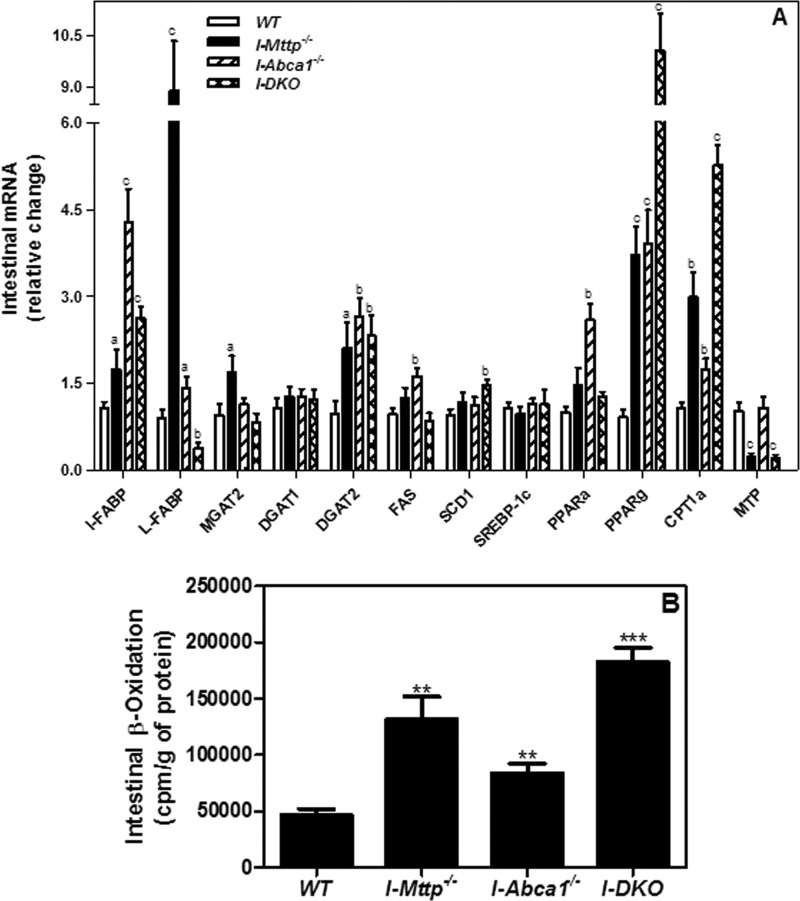
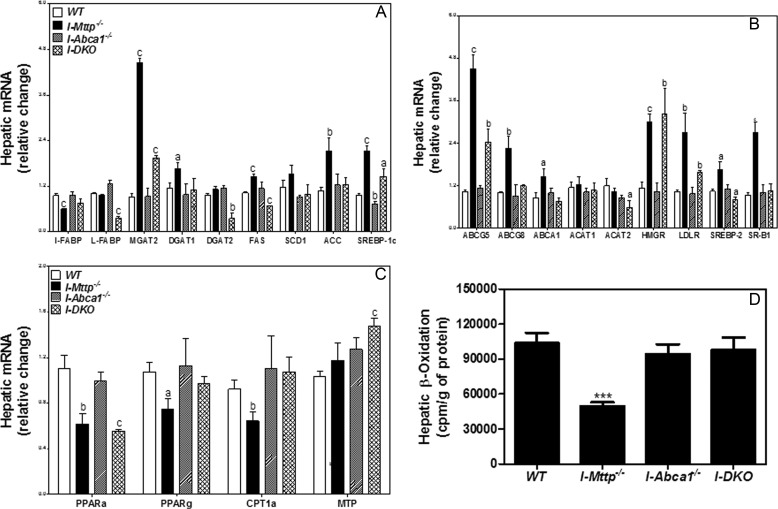
We have previously described apolipoprotein B (apoB)-dependent and -independent cholesterol absorption pathways and the role of microsomal triglyceride transfer protein (MTP) and ATP-binding cassette transporter A1 (ABCA1) in these pathways. To assess the contribution of these pathways to cholesterol absorption and to determine whether there are other pathways, we generated mice that lack MTP and ABCA1, individually and in combination, in the intestine. Intestinal deletions of Mttp and Abca1 decreased plasma cholesterol concentrations by 45 and 24%, respectively, whereas their combined deletion reduced it by 59%. Acute cholesterol absorption was reduced by 28% in the absence of ABCA1, and it was reduced by 92-95% when MTP was deleted in the intestine alone or together with ABCA1. MTP deficiency significantly reduced triglyceride absorption, although ABCA1 deficiency had no effect. ABCA1 deficiency did not affect cellular lipids, but Mttp deficiency significantly increased intestinal levels of triglycerides and free fatty acids. Accumulation of intestinal free fatty acids, but not triglycerides, in Mttp-deficient intestines was prevented when mice were also deficient in intestinal ABCA1. Combined deficiency of these genes increased intestinal fatty acid oxidation as a consequence of increased expression of peroxisome proliferator-activated receptor-γ (PPARγ) and carnitine palmitoyltransferase 1α (CPT1α). These studies show that intestinal MTP and ABCA1 are critical for lipid absorption and are the main determinants of plasma and intestinal lipid levels. Reducing their activities might lower plasma lipid concentrations.
Keywords: ABCA1; Cholesterol; Intestine; Lipid Absorption; Lipid Metabolism; Lipids; Lipoprotein Secretion; Lipoprotein Structure; MTP; Triglyceride.
Figures
References
-
- Grundy S. M., Cleeman J. I., Merz C. N., Brewer H. B., Jr., Clark L. T., Hunninghake D. B., Pasternak R. C., Smith S. C., Jr., Stone N. J. (2004) Implications of recent clinical trials for the National Cholesterol Education Program Adult Treatment Panel III Guidelines. Circulation 110, 227–239 - PubMed
-
- Berriot-Varoqueaux N., Aggerbeck L. P., Samson-Bouma M., Wetterau J. R. (2000) The role of the microsomal triglyceride transfer protein in abetalipoproteinemia. Annu. Rev. Nutr. 20, 663–697 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
