

Differential proliferation rates generate patterns of mechanical tension that orient tissue growth
- PMID: 24022370
- PMCID: PMC3817460
- DOI: 10.1038/emboj.2013.197
Differential proliferation rates generate patterns of mechanical tension that orient tissue growth
Abstract
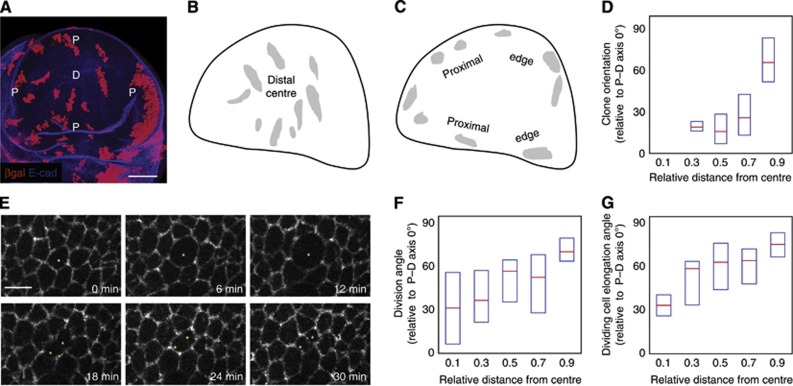
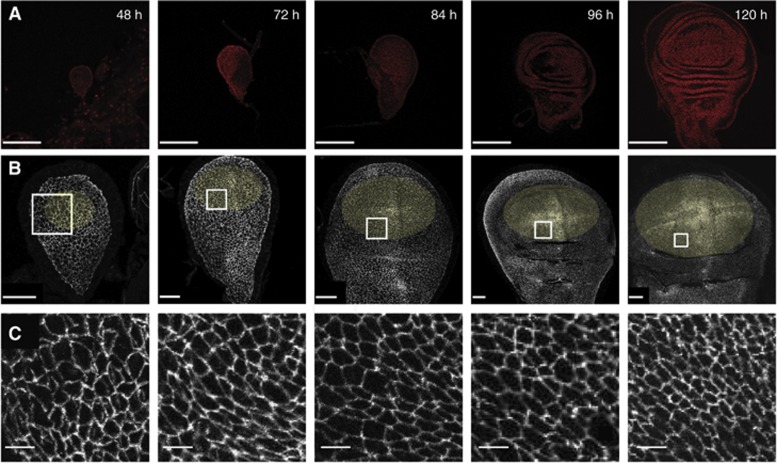
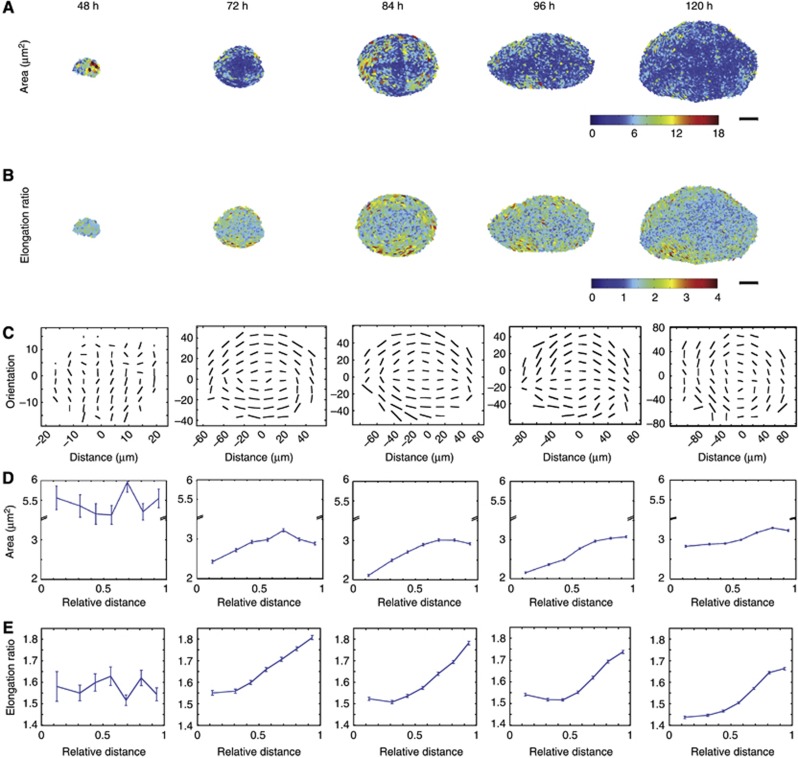
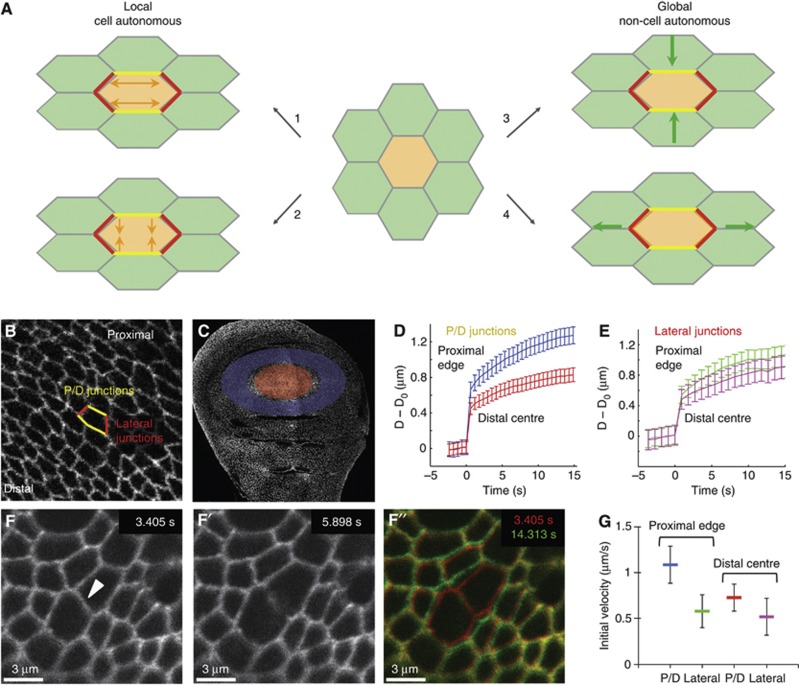
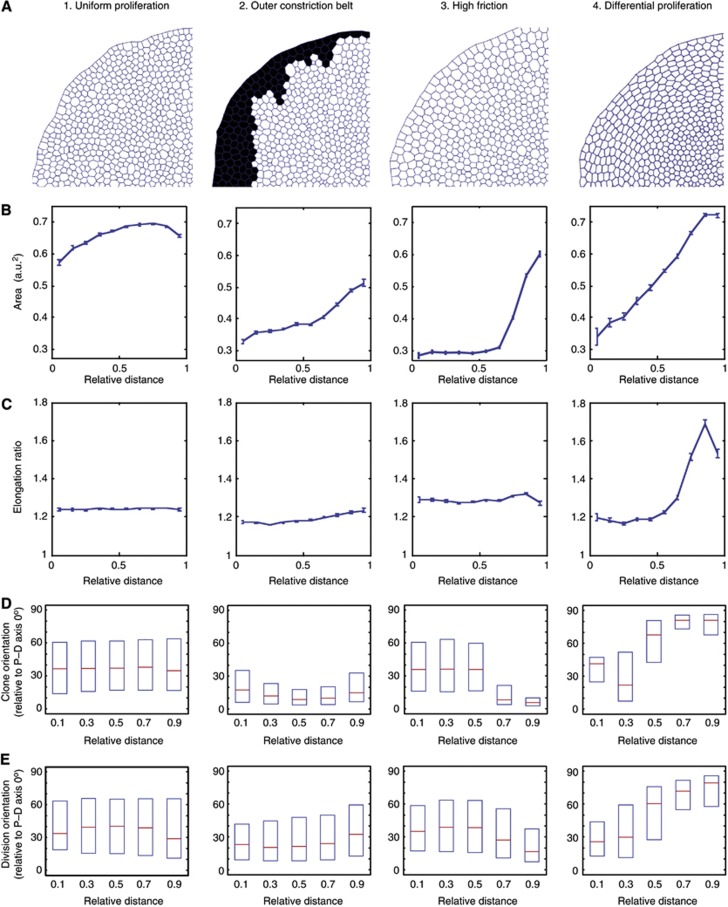
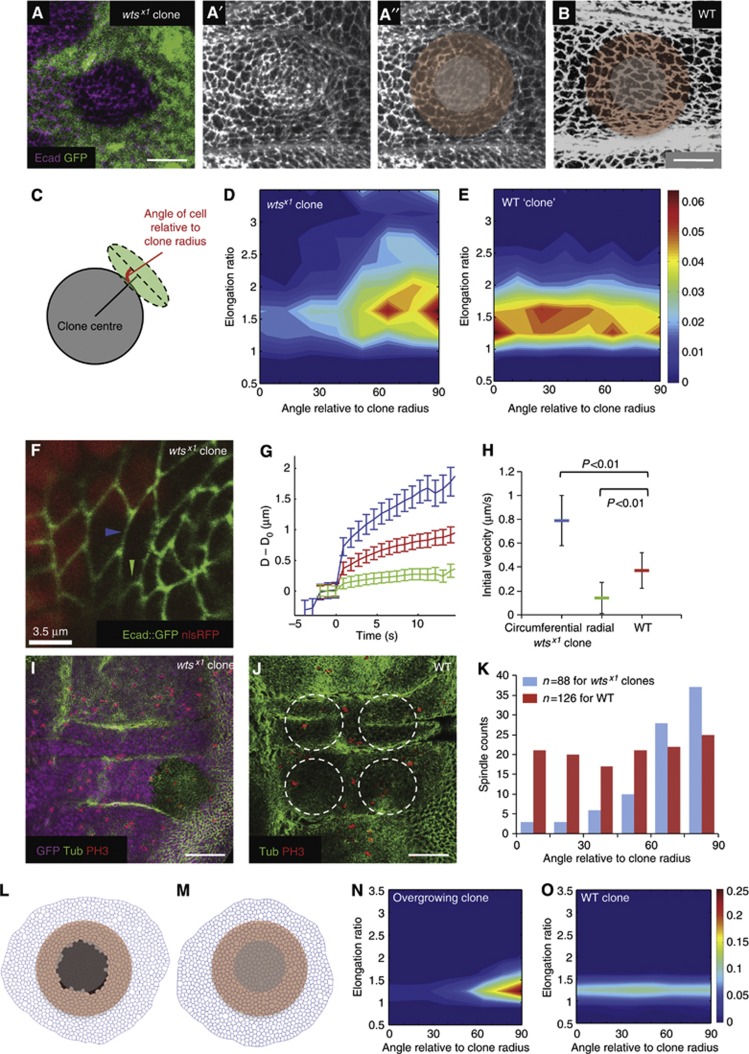
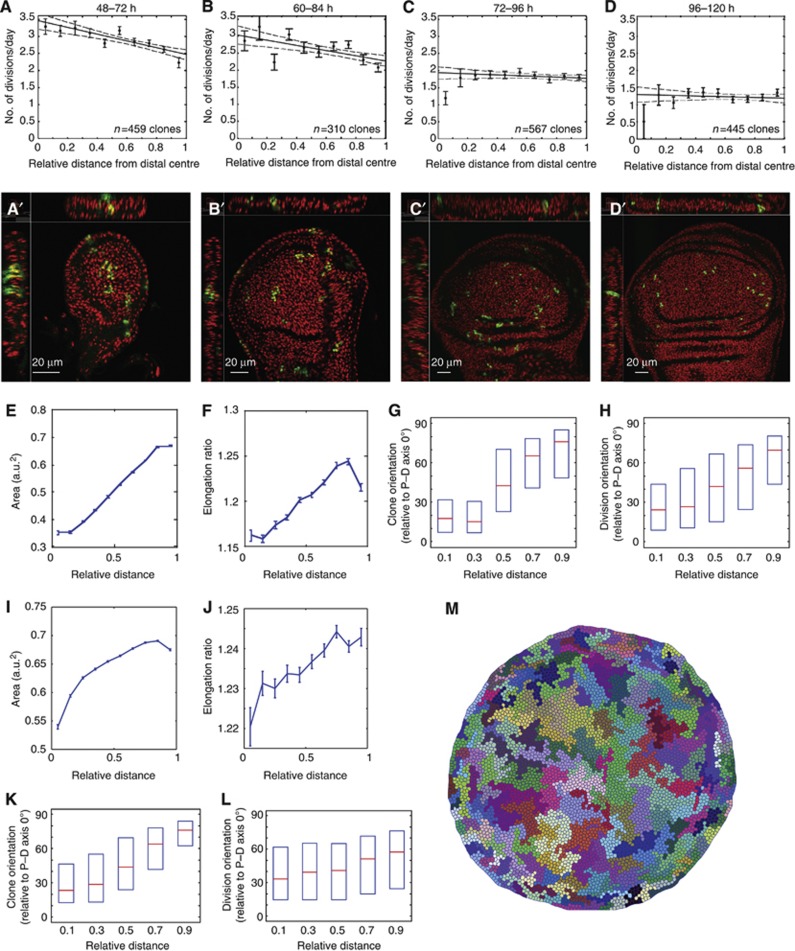
Orientation of cell divisions is a key mechanism of tissue morphogenesis. In the growing Drosophila wing imaginal disc epithelium, most of the cell divisions in the central wing pouch are oriented along the proximal-distal (P-D) axis by the Dachsous-Fat-Dachs planar polarity pathway. However, cells at the periphery of the wing pouch instead tend to orient their divisions perpendicular to the P-D axis despite strong Dachs polarization. Here, we show that these circumferential divisions are oriented by circumferential mechanical forces that influence cell shapes and thus orient the mitotic spindle. We propose that this circumferential pattern of force is not generated locally by polarized constriction of individual epithelial cells. Instead, these forces emerge as a global tension pattern that appears to originate from differential rates of cell proliferation within the wing pouch. Accordingly, we show that localized overgrowth is sufficient to induce neighbouring cell stretching and reorientation of cell division. Our results suggest that patterned rates of cell proliferation can influence tissue mechanics and thus determine the orientation of cell divisions and tissue shape.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
Comment in
-
The force and effect of cell proliferation.EMBO J. 2013 Oct 30;32(21):2783-4. doi: 10.1038/emboj.2013.225. Epub 2013 Oct 4. EMBO J. 2013. PMID: 24097062 Free PMC article.
References
-
- Aegerter-Wilmsen T, Aegerter CM, Hafen E, Basler K (2007) Model for the regulation of size in the wing imaginal disc of Drosophila. Mech Dev 124: 318–326 - PubMed
-
- Aegerter-Wilmsen T, Heimlicher MB, Smith AC, de Reuille PB, Smith RS, Aegerter CM, Basler K (2012) Integrating force-sensing and signaling pathways in a model for the regulation of wing imaginal disc size. Development 139: 3221–3231 - PubMed
-
- Aegerter-Wilmsen T, Smith AC, Christen AJ, Aegerter CM, Hafen E, Basler K (2010) Exploring the effects of mechanical feedback on epithelial topology. Development 137: 499–506 - PubMed
-
- Aigouy B, Farhadifar R, Staple DB, Sagner A, Röper JC, Jülicher F, Eaton S (2010) Cell flow reorients the axis of planar polarity in the wing epithelium of Drosophila. Cell 142: 773–786 - PubMed
-
- Aliee M, Roper JC, Landsberg KP, Pentzold C, Widmann TJ, Julicher F, Dahmann C (2012) Physical mechanisms shaping the Drosophila dorsoventral compartment boundary. Curr Biol 22: 967–976 - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
