

Review
doi: 10.1146/annurev-micro-092412-155620.
Wall teichoic acids of gram-positive bacteria
Affiliations
- PMID: 24024634
- PMCID: PMC3883102
- DOI: 10.1146/annurev-micro-092412-155620
Item in Clipboard
Review
Wall teichoic acids of gram-positive bacteria
Annu Rev Microbiol.
2013.
Abstract
The peptidoglycan layers of many gram-positive bacteria are densely functionalized with anionic glycopolymers known as wall teichoic acids (WTAs). These polymers play crucial roles in cell shape determination, regulation of cell division, and other fundamental aspects of gram-positive bacterial physiology. Additionally, WTAs are important in pathogenesis and play key roles in antibiotic resistance. We provide an overview of WTA structure and biosynthesis, review recent studies on the biological roles of these polymers, and highlight remaining questions. We also discuss prospects for exploiting WTA biosynthesis as a target for new therapies to overcome resistant infections.
Figures
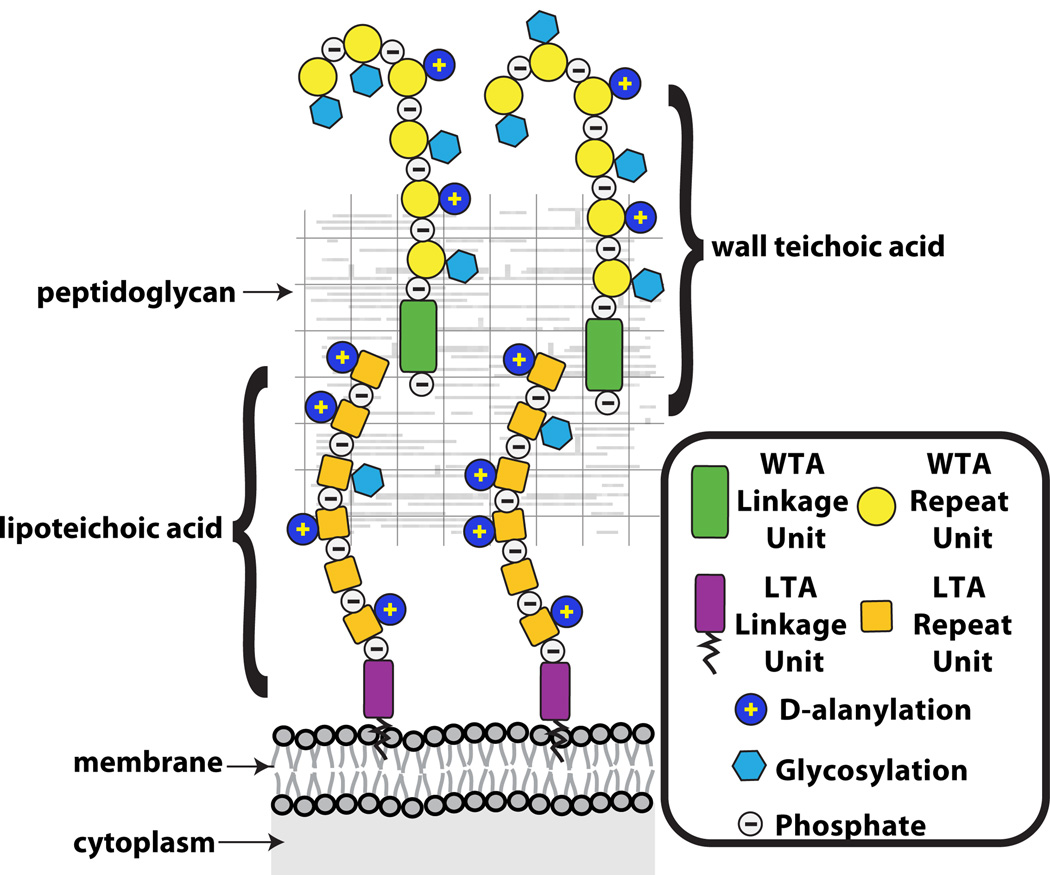
Schematic of the gram-positive cell wall showing that wall teichoic acids are covalently anchored to peptidoglycan and lipoteichoic acids are tethered to the membrane. The WTAs extend beyond the PG layer, whereas fully extended LTAs may not be able to reach pass the PG layer.
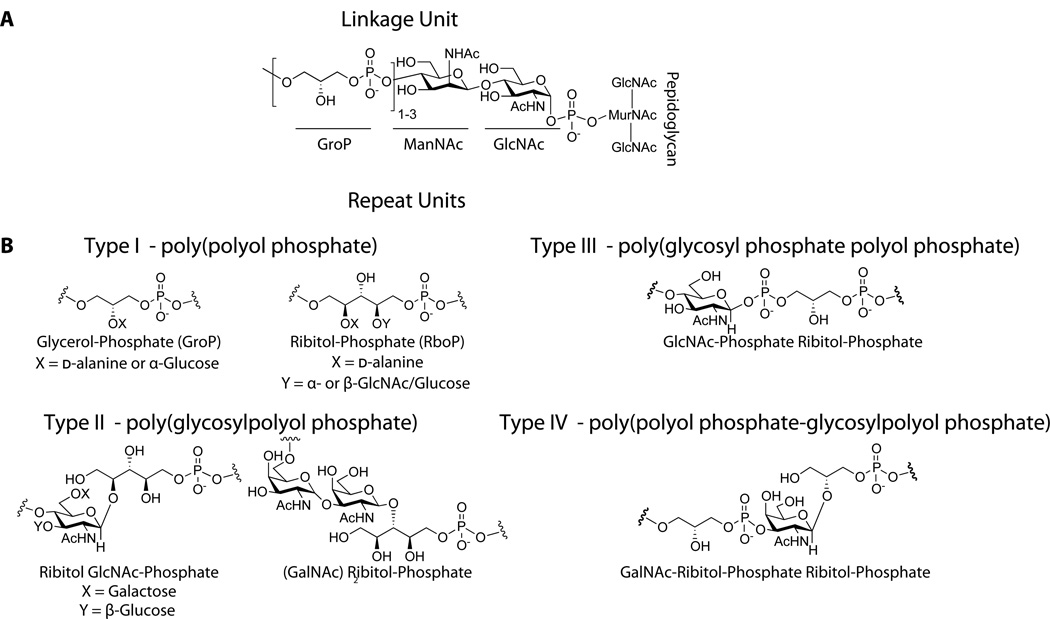
A. The commonly characterized WTA linkage unit consists of a GroP-ManNAc-GlcNAc-Phosphate that is covalently attached to peptidoglycan. While the stereochemistry of the glycosidic linkage has not been clearly established, it is assumed to be retained as α based on predicted enzymatic catalysis. B. Four different classes of WTA repeat units found in gram-positive bacteria (90).
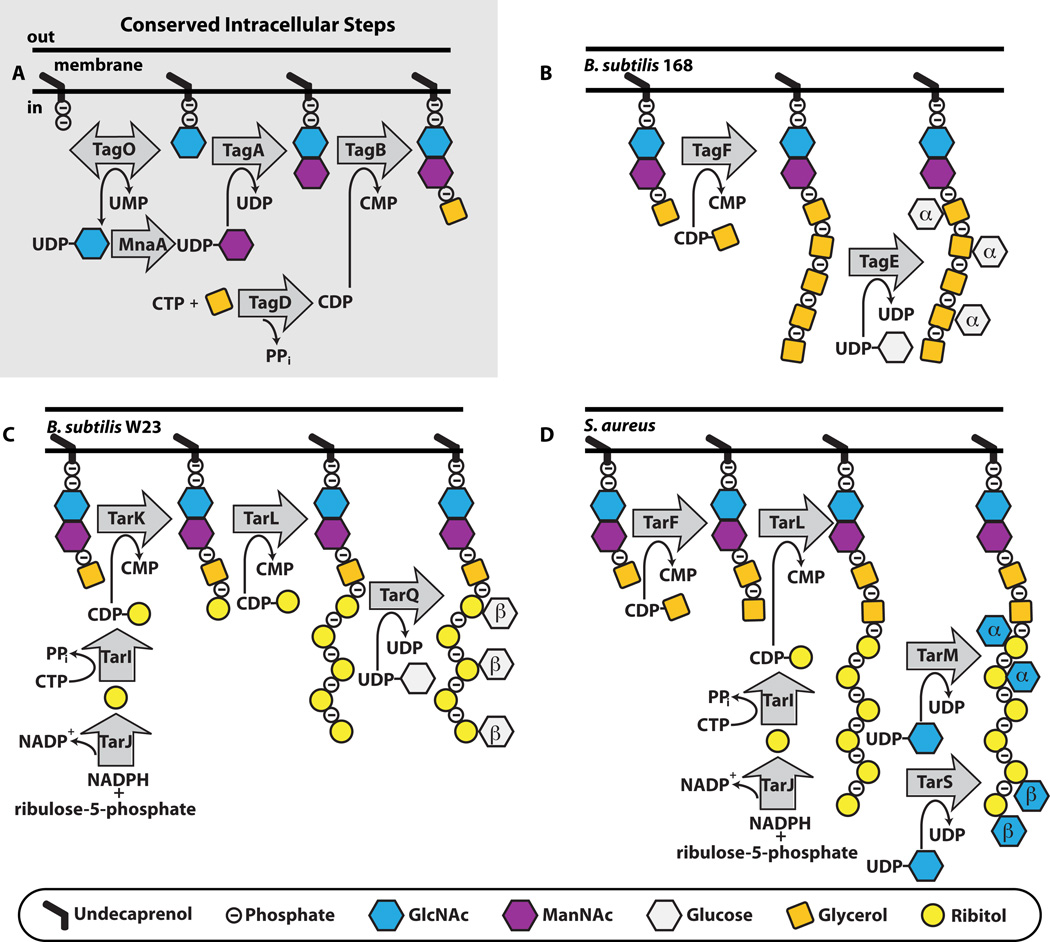
A. The initial steps to WTA polymer formation that has been found in all thus far characterized pathways. B,C,D. The end stage intracellular steps to form the WTA polymer in B. subtilis 168 (B), B. subtilis W23 (C) and S. aureus (D). TarIJ reactions to form CDP-ribitol exist in both B. subtilis W23 and S. aureus. The α- and β- within the sugar denotes the stereochemistry of attachment. We have drawn α- and β-GlcNAc attachment to S. aureus occurring on the same strand, but it is possible that the TarS and TarM enzymes do not attach GlcNAc to the same polymer.
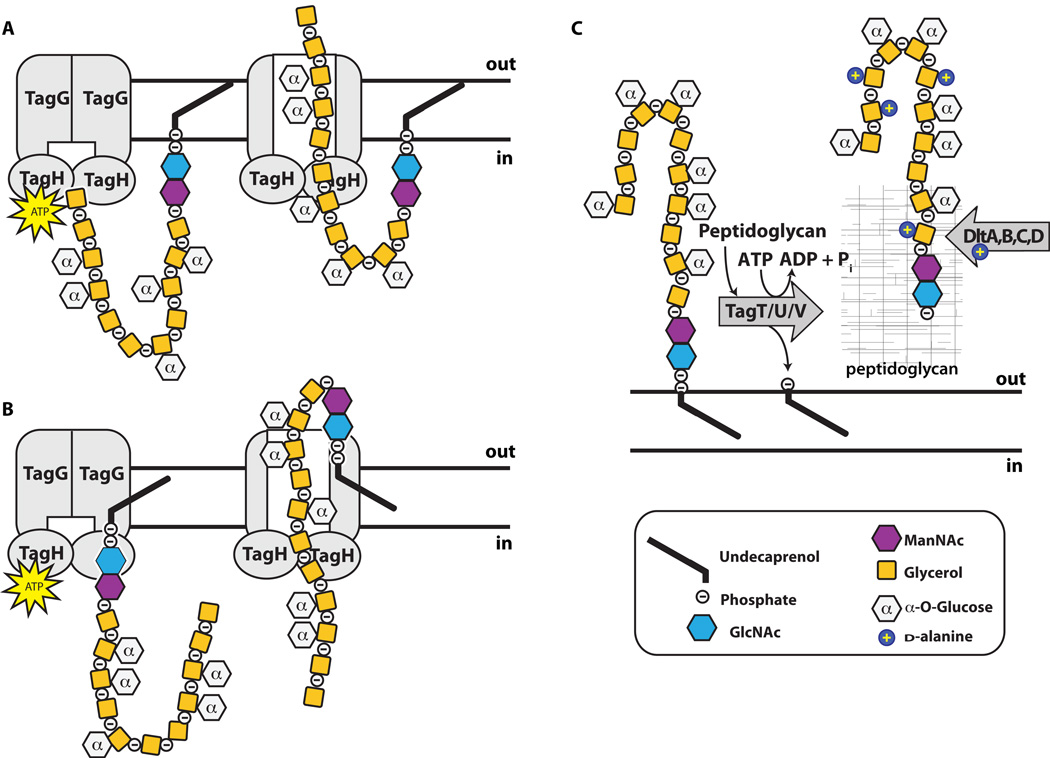
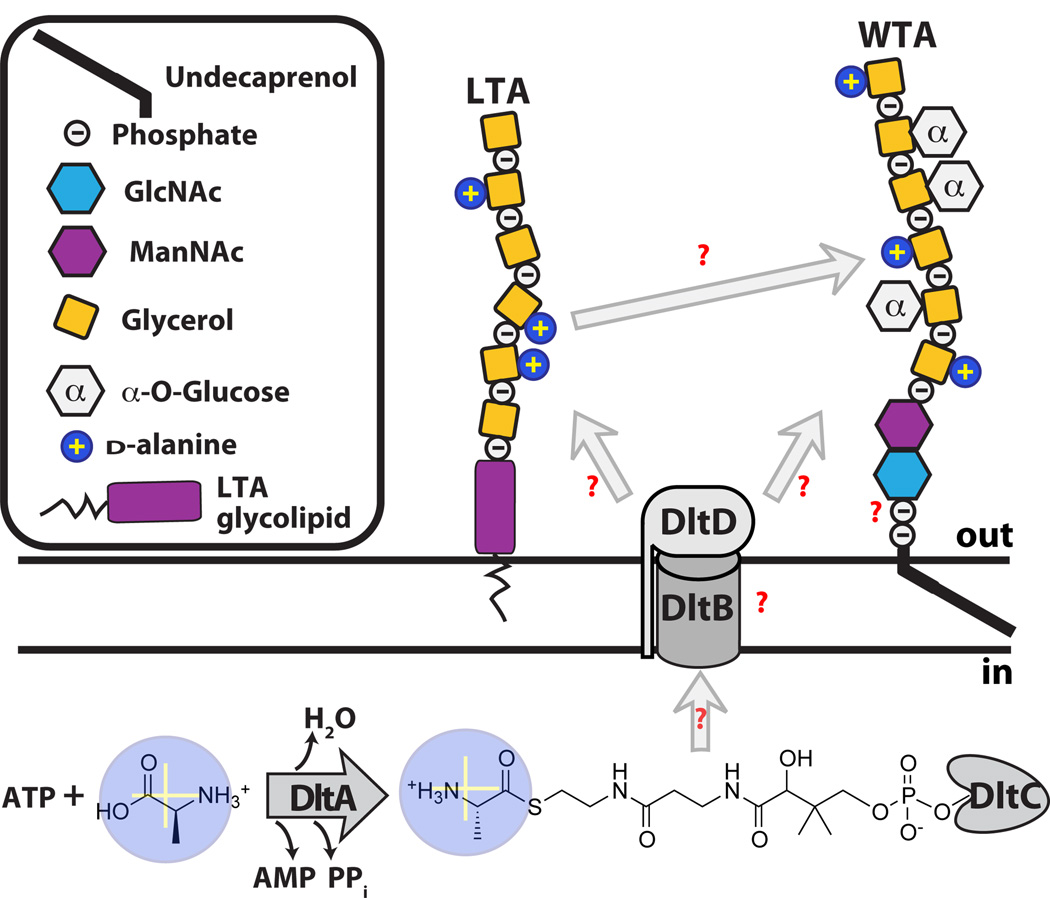
Graphical representation of the poly(GroP) WTA polymer in B. subtilis 168 transported through the cell membrane by TagGH. Homology suggests energy generated by TagH ATP hydrolysis drives a conformational change that allows for polymer transport. TagGH can presumably either thread the polymer through the membrane by the non-reducing end (A) or recognize the disaccharide linkage unit and flip the polymer across (B). Following transport (C), the polymer is covalently attached to PG by TagTUV. It is unknown whether the TagTUV enzymes work independently or together to ligate WTA to PG. The presumed energy source for this ligation reaction, which we designated as ATP hydrolysis, is undetermined. It remains unknown whether extracellular d -alanylation by DltABCD occurs before or after WTA is covalently attached to PG.
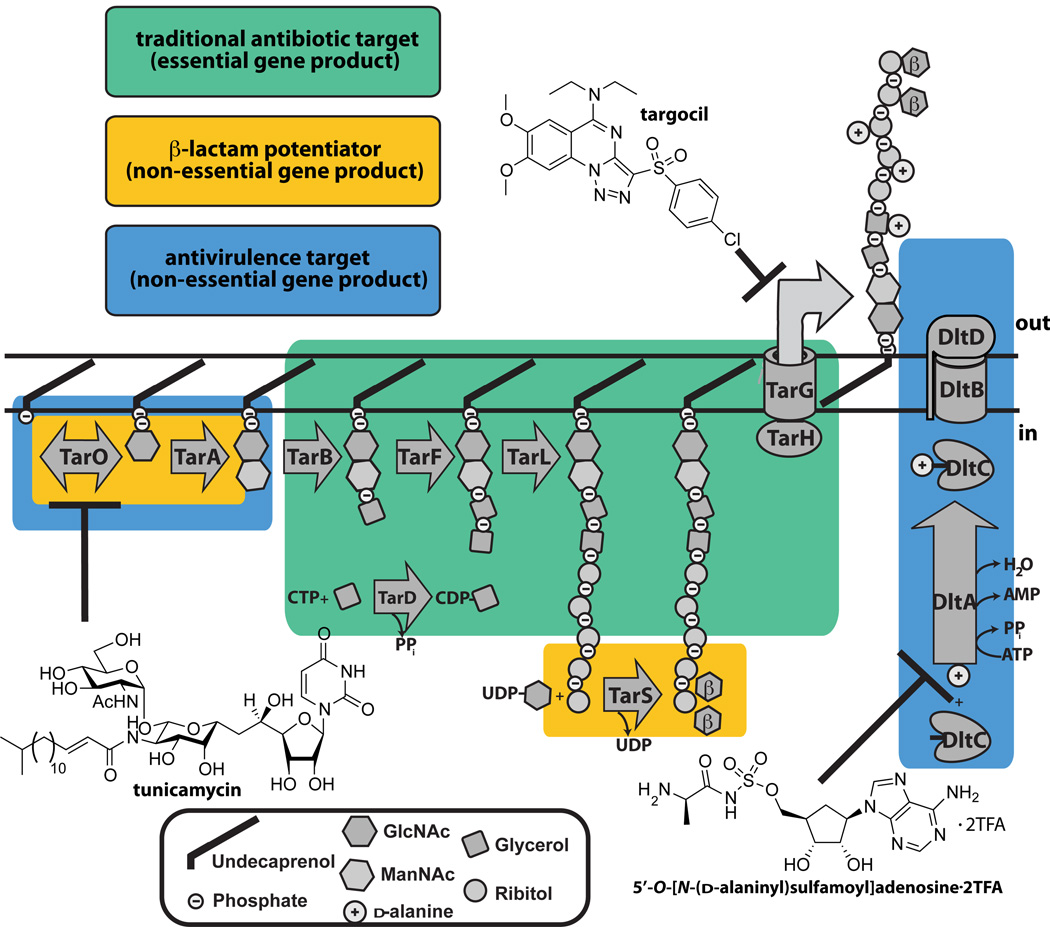
Schematic showing the three types of antibacterial targets present in the S. aureus WTA pathway. Enzymes that are traditional, β-lactam potentiators, or antivirulence antimicrobial targets are boxed. The three chemical structures illustrated are small molecules that are known to inhibit the WTA enzymes, TarO, TarG, and DltA.
References
-
- Aly R, Shinefield HR, Litz C, Maibach HI. Role of teichoic acid in the binding of Staphylococcus aureus to nasal epithelial cells. J Infect Dis. 1980;141:463–465. - PubMed
-
- Anderson AJ, Green RS, Sturman AJ, Archibald AR. Cell wall assembly in Bacillus subtilis: location of wall material incorporated during pulsed release of phosphate limitation, its accessibility to bacteriophages and concanavalin A, and its susceptibility to turnover. J Bacteriol. 1978;136:886–899. - PMC - PubMed
-
- Andre G, Deghorain M, Bron PA, van S, II, Kleerebezem M, et al. Fluorescence and atomic force microscopy imaging of wall teichoic acids in Lactobacillus plantarum. ACS Chem Biol. 2011;6:366–376. - PubMed
-
- Araki Y, Ito E. Linkage units in cell walls of gram-positive bacteria. Crit Rev Microbiol. 1989;17:121–135. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
