

An ENU mutagenesis screen identifies novel and known genes involved in epigenetic processes in the mouse
- PMID: 24025402
- PMCID: PMC4053835
- DOI: 10.1186/gb-2013-14-9-r96
An ENU mutagenesis screen identifies novel and known genes involved in epigenetic processes in the mouse
Abstract
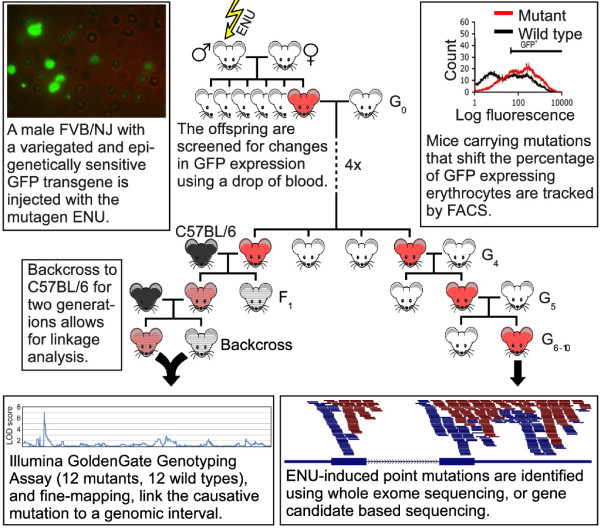
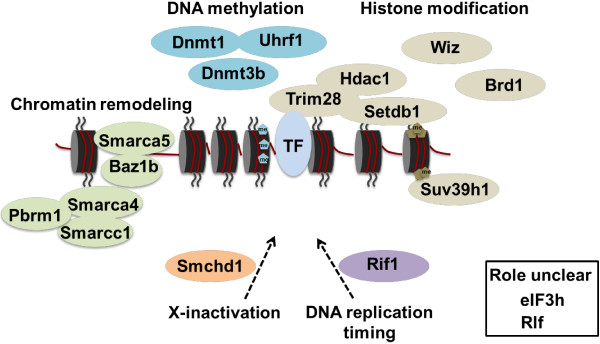
Background: We have used a sensitized ENU mutagenesis screen to produce mouse lines that carry mutations in genes required for epigenetic regulation. We call these lines Modifiers of murine metastable epialleles (Mommes).
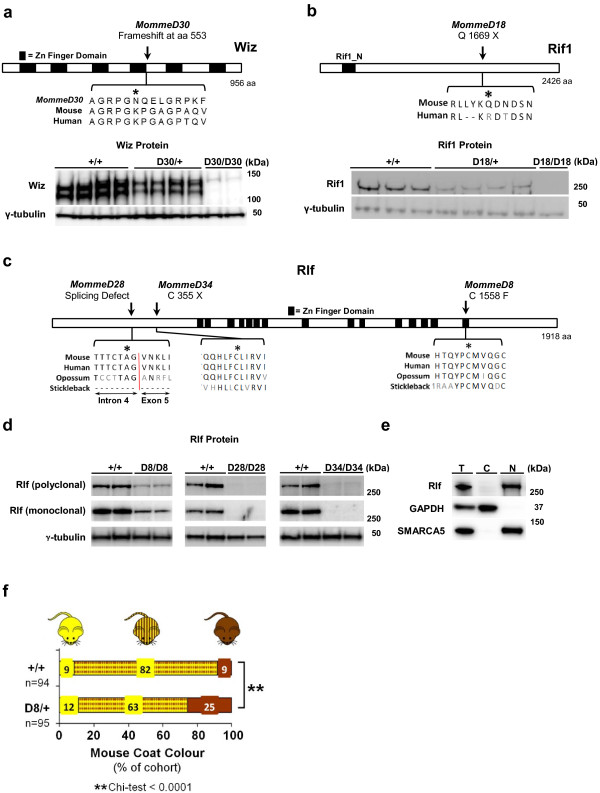
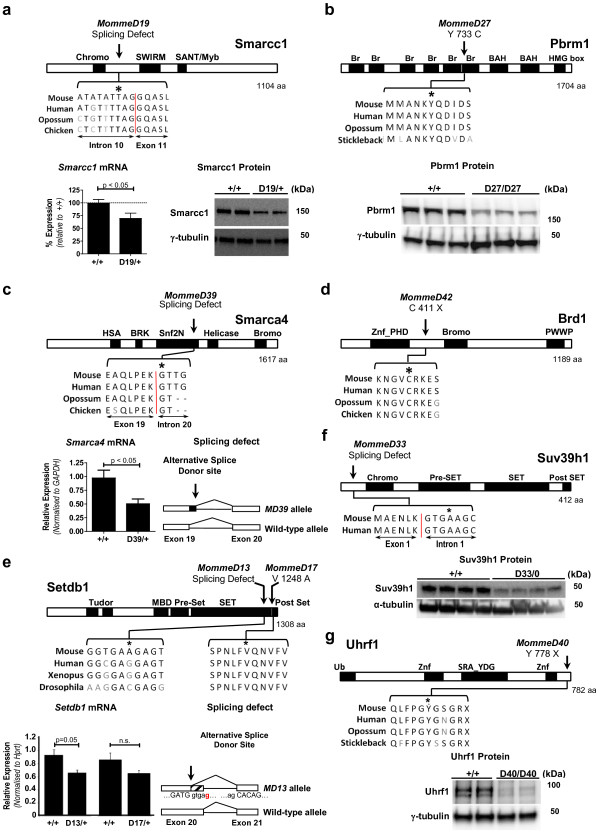
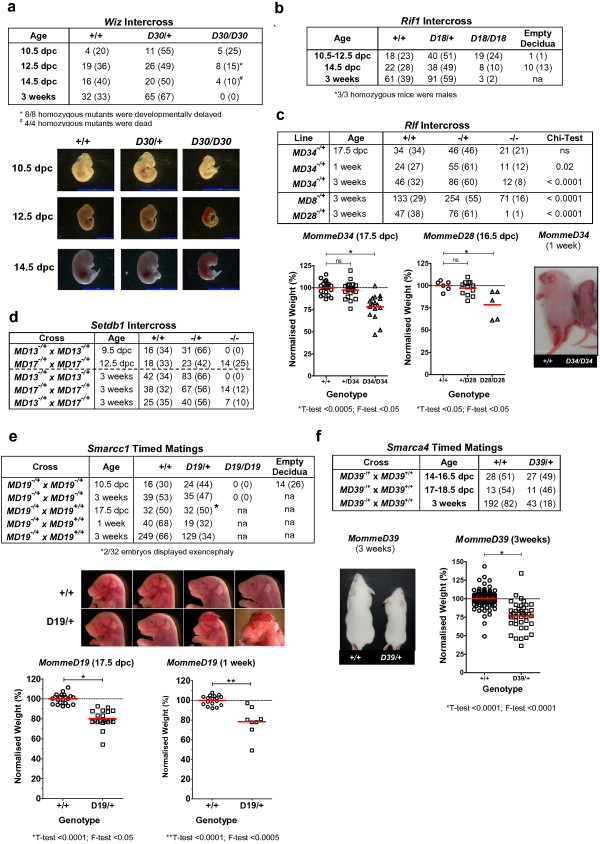
Results: We report a basic molecular and phenotypic characterization for twenty of the Momme mouse lines, and in each case we also identify the causative mutation. Three of the lines carry a mutation in a novel epigenetic modifier, Rearranged L-myc fusion (Rlf), and one gene, Rap-interacting factor 1 (Rif1), has not previously been reported to be involved in transcriptional regulation in mammals. Many of the other lines are novel alleles of known epigenetic regulators. For two genes, Rlf and Widely-interspaced zinc finger (Wiz), we describe the first mouse mutants. All of the Momme mutants show some degree of homozygous embryonic lethality, emphasizing the importance of epigenetic processes. The penetrance of lethality is incomplete in a number of cases. Similarly ,abnormalities in phenotype seen in the heterozygous individuals of some lines occur with incomplete penetrance.
Conclusions: Recent advances in sequencing enhance the power of sensitized mutagenesis screens to identify the function of previously uncharacterized factors and to discover additional functions for previously characterized proteins. The observation of incomplete penetrance of phenotypes in these inbred mutant mice, at various stages of development, is of interest. Overall, the Momme collection of mouse mutants provides a valuable resource for researchers across many disciplines.
Figures

References
-
- Henikoff S. Position-effect variegation after 60 years. Trends Genet. 1990;14:422–426. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
