

Congenital hypoplasia of the cerebellum: developmental causes and behavioral consequences
- PMID: 24027500
- PMCID: PMC3759752
- DOI: 10.3389/fnana.2013.00029
Congenital hypoplasia of the cerebellum: developmental causes and behavioral consequences
Abstract
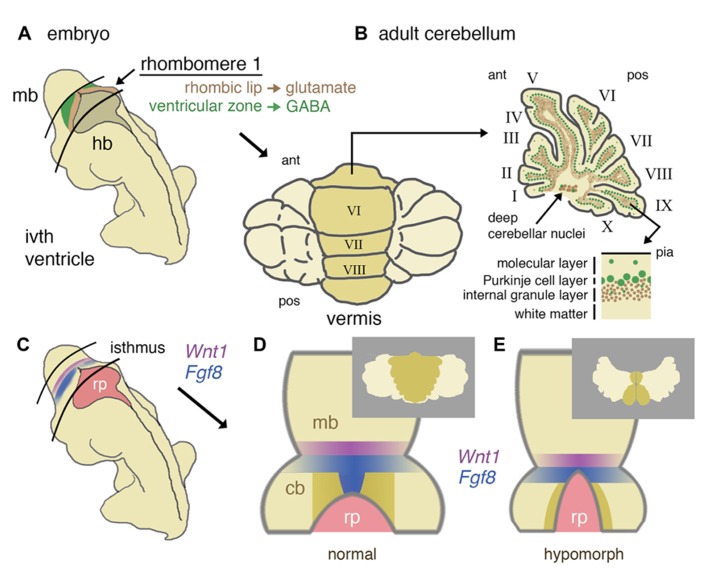
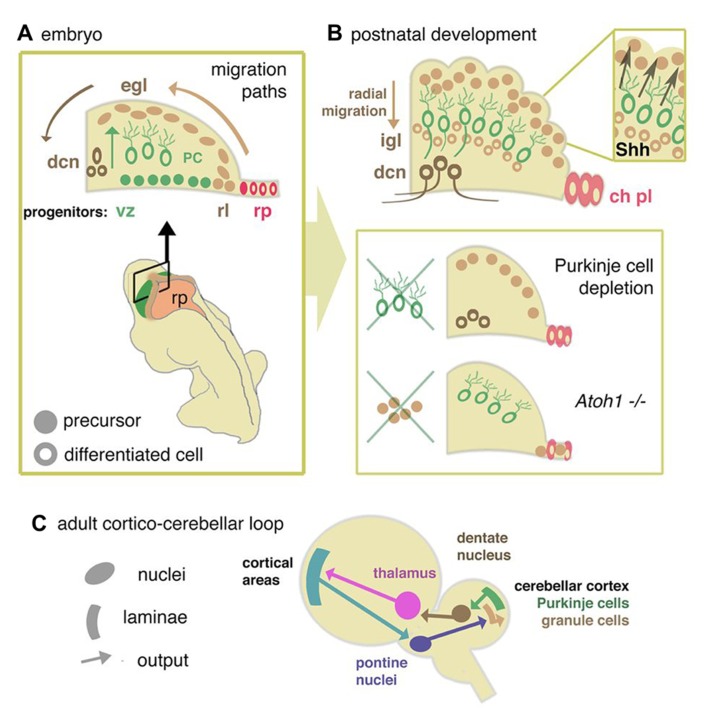
Over the last 60 years, the spotlight of research has periodically returned to the cerebellum as new techniques and insights have emerged. Because of its simple homogeneous structure, limited diversity of cell types and characteristic behavioral pathologies, the cerebellum is a natural home for studies of cell specification, patterning, and neuronal migration. However, recent evidence has extended the traditional range of perceived cerebellar function to include modulation of cognitive processes and implicated cerebellar hypoplasia and Purkinje neuron hypo-cellularity with autistic spectrum disorder. In the light of this emerging frontier, we review the key stages and genetic mechanisms behind cerebellum development. In particular, we discuss the role of the midbrain hindbrain isthmic organizer in the development of the cerebellar vermis and the specification and differentiation of Purkinje cells and granule neurons. These developmental processes are then considered in relation to recent insights into selected human developmental cerebellar defects: Joubert syndrome, Dandy-Walker malformation, and pontocerebellar hypoplasia. Finally, we review current research that opens up the possibility of using the mouse as a genetic model to study the role of the cerebellum in cognitive function.
Keywords: autism spectrum disorders; behavior; cerebellum; defects; development; function; genetics; hypoplasia.
Figures
Similar articles
-
Cerebellar development and disease.Curr Opin Neurobiol. 2008 Feb;18(1):12-9. doi: 10.1016/j.conb.2008.05.010. Epub 2008 May 29. Curr Opin Neurobiol. 2008. PMID: 18513948 Free PMC article. Review.
-
Recessive PRDM13 mutations cause fatal perinatal brainstem dysfunction with cerebellar hypoplasia and disrupt Purkinje cell differentiation.Am J Hum Genet. 2022 May 5;109(5):909-927. doi: 10.1016/j.ajhg.2022.03.010. Epub 2022 Apr 6. Am J Hum Genet. 2022. PMID: 35390279 Free PMC article.
-
Human malformations of the midbrain and hindbrain: review and proposed classification scheme.Mol Genet Metab. 2003 Sep-Oct;80(1-2):36-53. doi: 10.1016/j.ymgme.2003.08.010. Mol Genet Metab. 2003. PMID: 14567956 Review.
-
Clinical nosologic and genetic aspects of Joubert and related syndromes.J Child Neurol. 1999 Oct;14(10):660-6; discussion 669-72. doi: 10.1177/088307389901401007. J Child Neurol. 1999. PMID: 10511339 Review.
-
Developmental outcomes in children with congenital cerebellar malformations.Dev Med Child Neurol. 2019 Mar;61(3):350-358. doi: 10.1111/dmcn.14059. Epub 2018 Oct 15. Dev Med Child Neurol. 2019. PMID: 30320441
Cited by
-
Components of Endocannabinoid Signaling System Are Expressed in the Perinatal Mouse Cerebellum and Required for Its Normal Development.eNeuro. 2020 Apr 23;7(2):ENEURO.0471-19.2020. doi: 10.1523/ENEURO.0471-19.2020. Print 2020 Mar/Apr. eNeuro. 2020. PMID: 32179579 Free PMC article.
-
A Randomized Controlled Trial of Sertraline in Young Children With Autism Spectrum Disorder.Front Psychiatry. 2019 Nov 6;10:810. doi: 10.3389/fpsyt.2019.00810. eCollection 2019. Front Psychiatry. 2019. PMID: 31780970 Free PMC article.
-
A loss-of-function mutation in human Oxidation Resistance 1 disrupts the spatial-temporal regulation of histone arginine methylation in neurodevelopment.Genome Biol. 2023 Sep 29;24(1):216. doi: 10.1186/s13059-023-03037-1. Genome Biol. 2023. PMID: 37773136 Free PMC article.
-
Distribution and Functional Implication of Secretin in Multiple Brain Regions.J Mol Neurosci. 2019 Jul;68(3):485-493. doi: 10.1007/s12031-018-1089-z. Epub 2018 Jun 7. J Mol Neurosci. 2019. PMID: 29882022 Review.
-
Fetal exposure to valproic acid dysregulates the expression of autism-linked genes in the developing cerebellum.Transl Psychiatry. 2023 Apr 5;13(1):114. doi: 10.1038/s41398-023-02391-9. Transl Psychiatry. 2023. PMID: 37019889 Free PMC article.
References
-
- Abdelhamed Z. A., Wheway G., Szymanska K., Natarajan S., Toomes C., Inglehearn C., et al. (2013). Variable expressivity of ciliopathy neurological phenotypes that encompass Meckel–Gruber syndrome and Joubert syndrome is caused by complex de-regulated ciliogenesis, Shh and Wnt signalling defects. Hum. Mol. Genet. 22 1358–137210.1093/hmg/dds546 - DOI - PMC - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources