

Chloride channel ClC-2 is a key factor in the development of DSS-induced murine colitis
- PMID: 24030525
- PMCID: PMC3880115
- DOI: 10.1097/MIB.0b013e3182a82ae9
Chloride channel ClC-2 is a key factor in the development of DSS-induced murine colitis
Abstract
Background: Previously, it was shown that the chloride channel ClC-2 modulates intestinal tight junction (TJ) barrier function. The aim of the present study was to investigate the role of ClC-2 in epithelial barrier function and recovery in the event of epithelial injury.
Methods: The role of ClC-2 was investigated in TJ barrier function in dextran sodium sulfate (DSS)-induced colitis in ClC-2 knockout mice and ClC-2 knockdown intestinal Caco-2 cells. Barrier function was measured electrophysiologically and by transepithelial mannitol fluxes. Selected TJ and associated proteins were Western blotted, cytokines were measured using quantitative PCR, and human colonic biopsies were examined with immunohistochemistry.
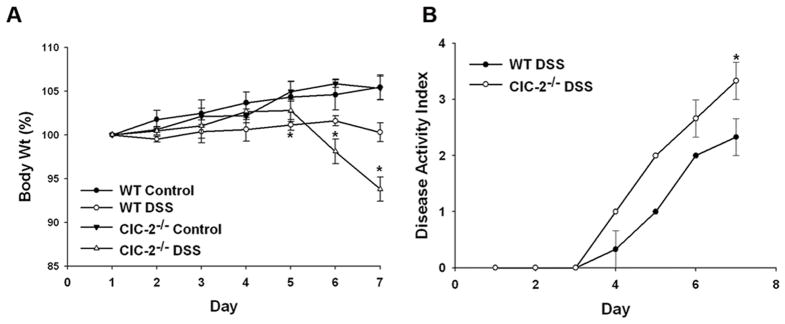
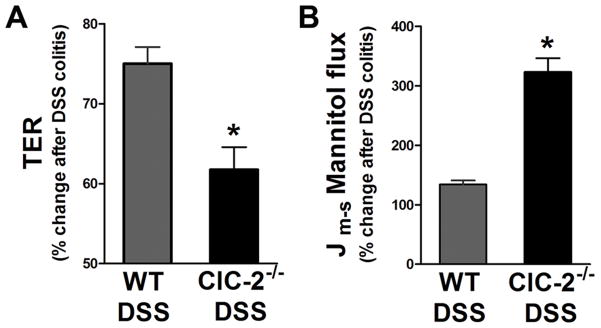
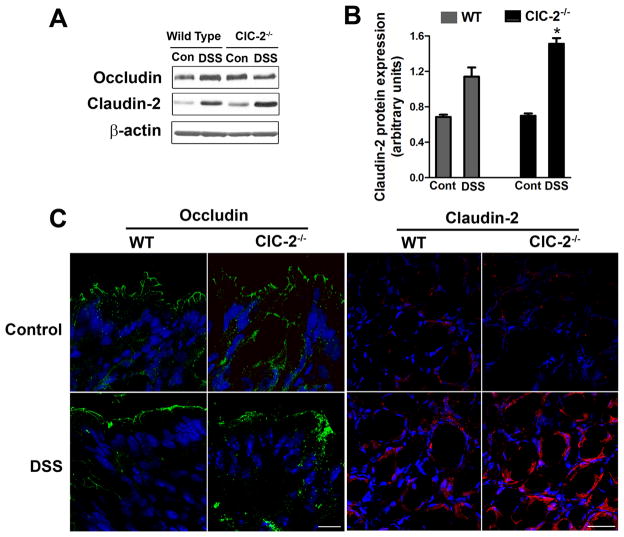
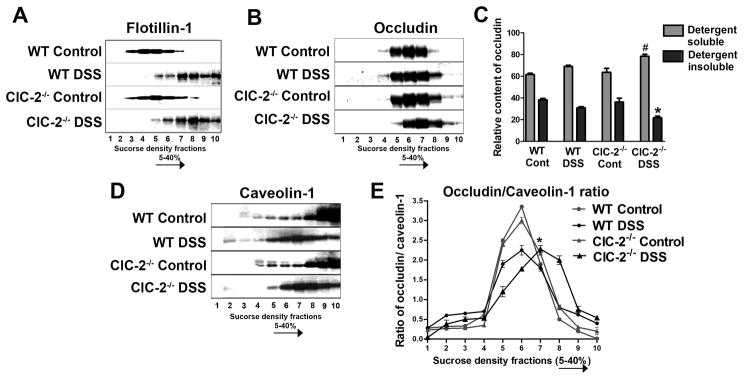
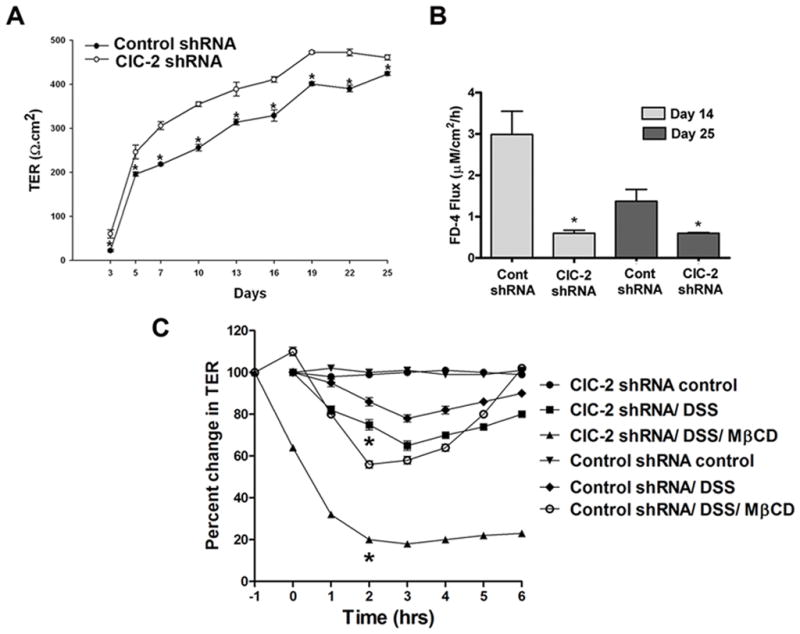
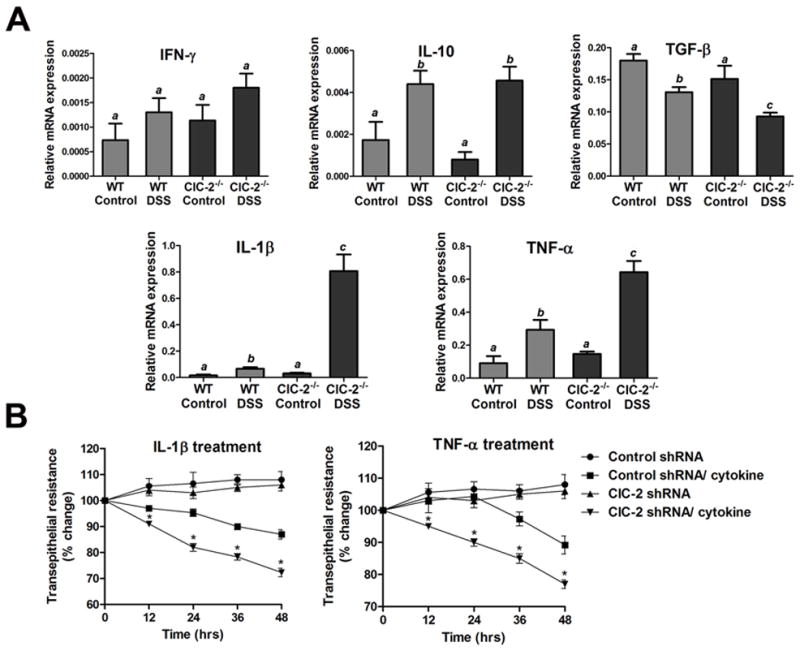
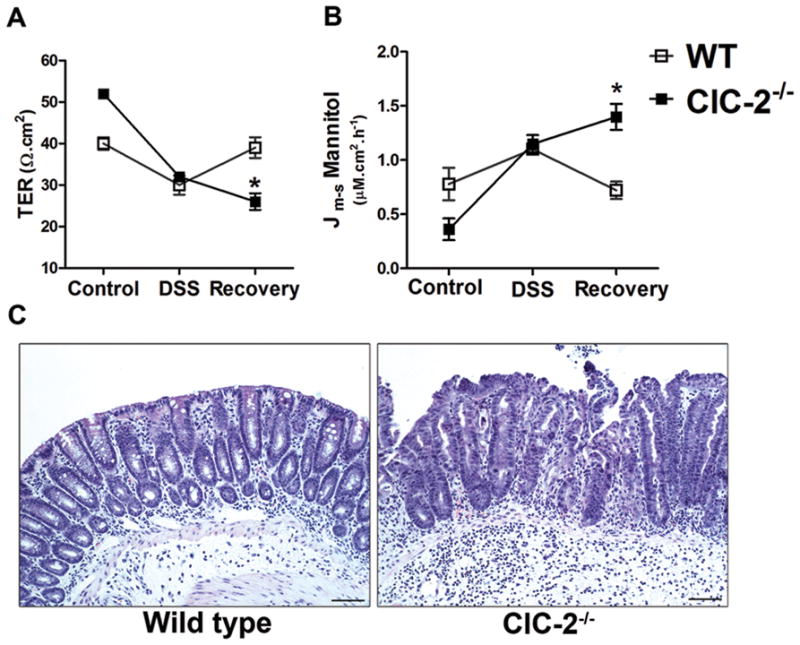
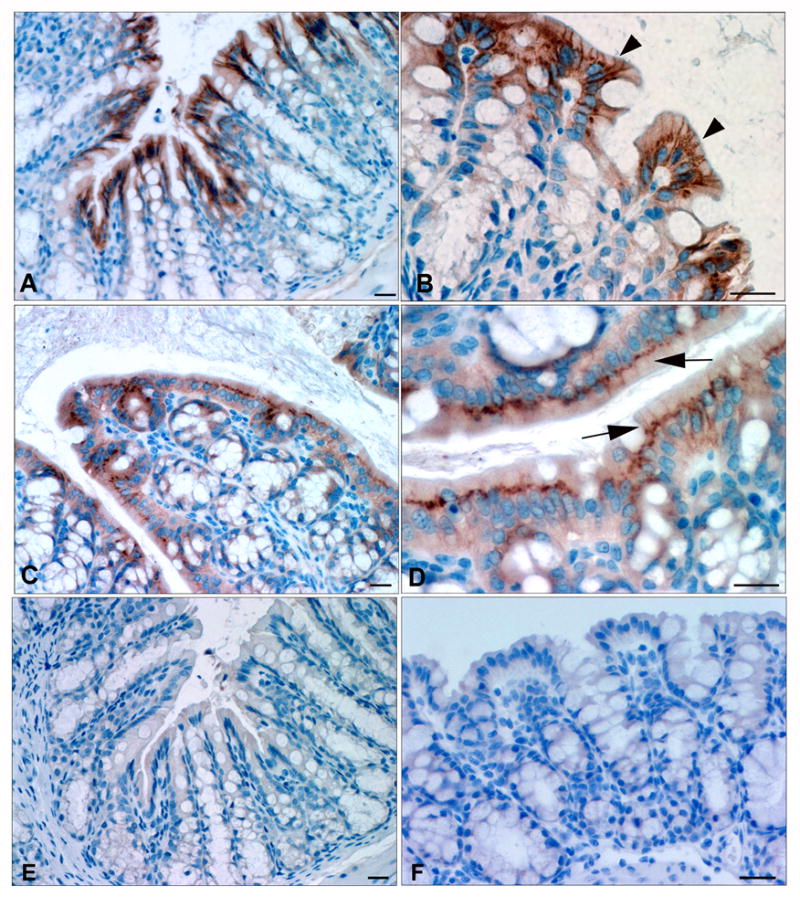
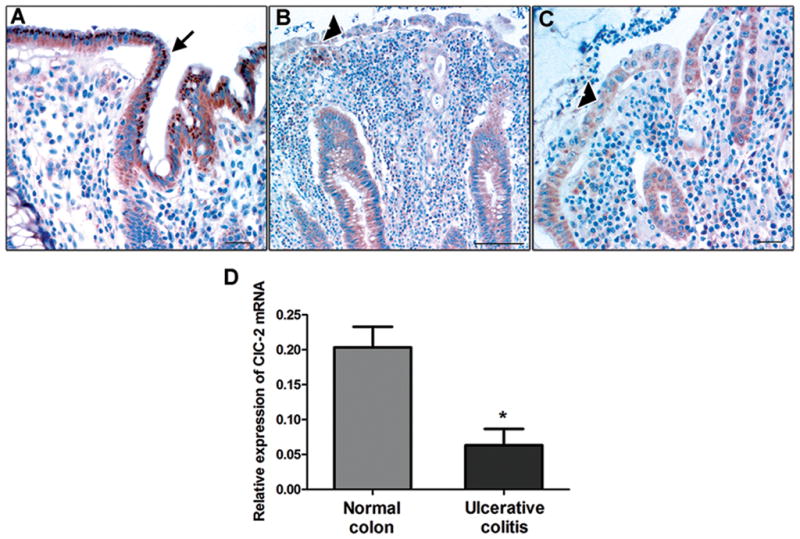
Results: ClC-2 mice had a higher disease activity index, higher histological scores, and increased paracellular permeability compared with wild-type mice when treated with DSS. DSS-treated ClC-2 mice had increased claudin-2 expression, greater loss of occludin in the membrane, increased association of occludin with caveolin-1, and significantly increased tumor necrosis factor-α and interleukin-1β messenger RNA. ClC-2 knockdown in human intestinal Caco-2 cells resulted in a greater loss of epithelial resistance in the event of epithelial injury. The restoration of colonic barrier function after DSS colitis was delayed in ClC-2 mice. In human colonic biopsies, the protein and messenger RNA expression of ClC-2 was found to be reduced in patients with ulcerative colitis.
Conclusions: ClC-2 plays a critical role in experimental colitis in that its absence increases disease activity, reduces barrier function and recovery, and perturbs TJs. Furthermore, ClC-2 expression is markedly reduced in the colon of human patients with ulcerative colitis.
Conflict of interest statement
Figures

References
-
- Nilius B, Droogmans G. Amazing chloride channels: an overview. Acta Physiol Scand. 2003;177:119–147. - PubMed
-
- Jentsch TJ. CLC chloride channels and transporters: from genes to protein structure, pathology and physiology. Crit Rev Biochem Mol Biol. 2008;43:3–36. - PubMed
-
- Niemeyer MI, Yusef YR, Cornejo I, et al. Functional evaluation of human ClC-2 chloride channel mutations associated with idiopathic generalized epilepsies. Physiol Genomics. 2004;19:74–83. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases