

Mutagenic analysis of the intracellular portals of the human 5-HT3A receptor
- PMID: 24030822
- PMCID: PMC3814755
- DOI: 10.1074/jbc.M113.503300
Mutagenic analysis of the intracellular portals of the human 5-HT3A receptor
Abstract
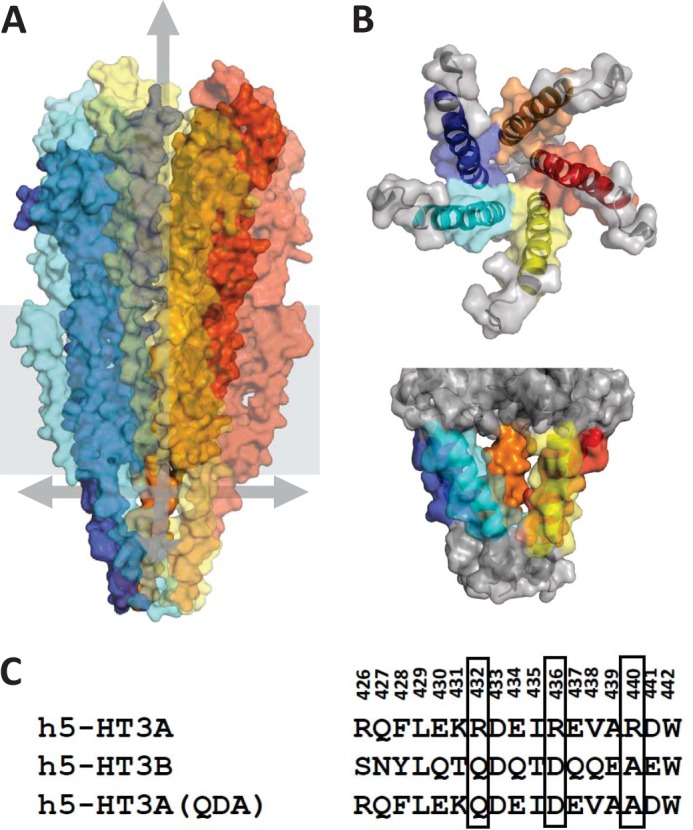
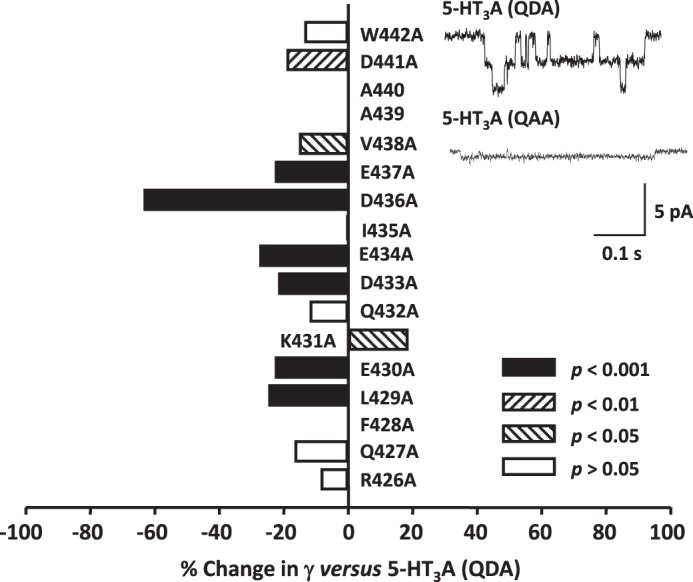
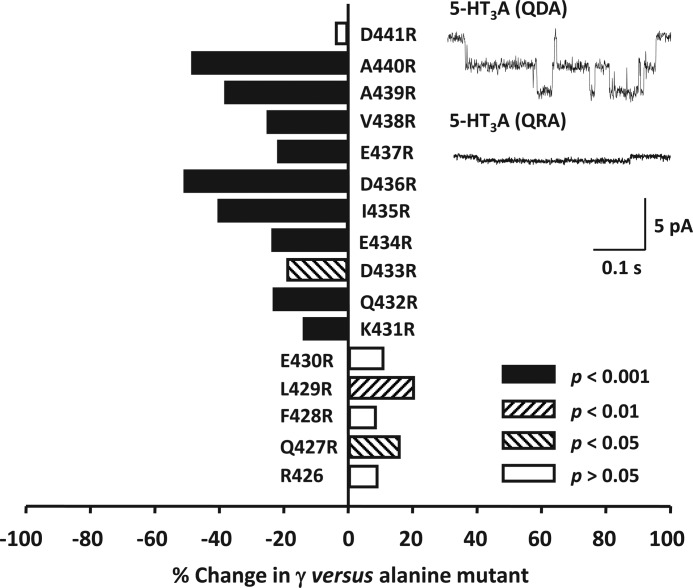
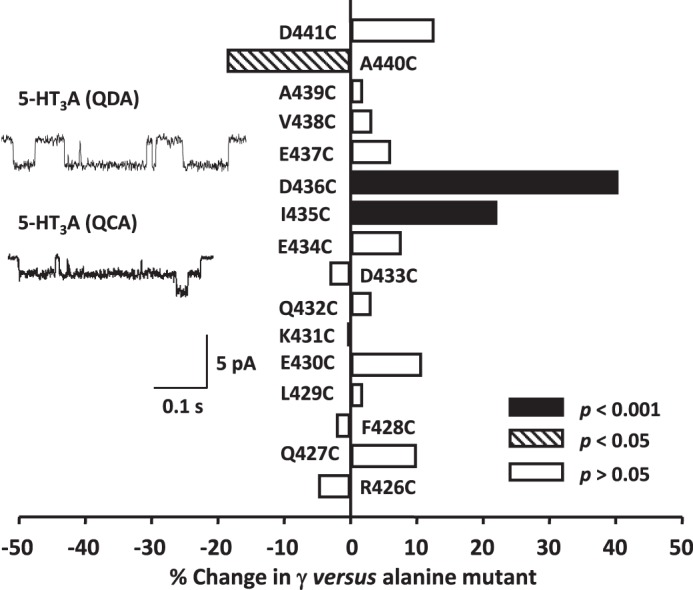
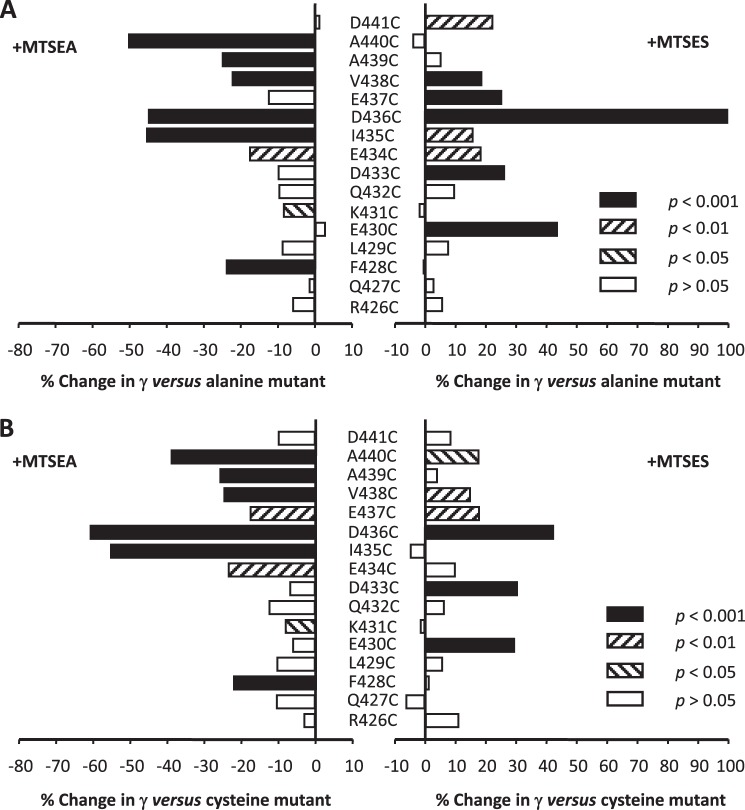
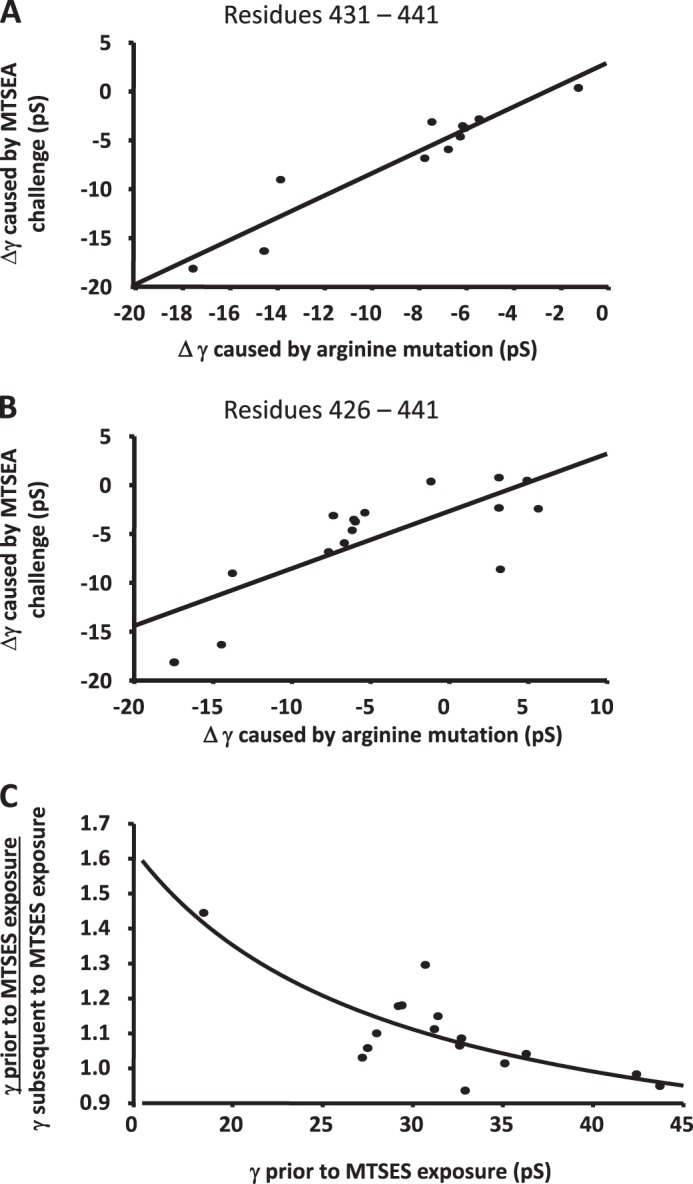
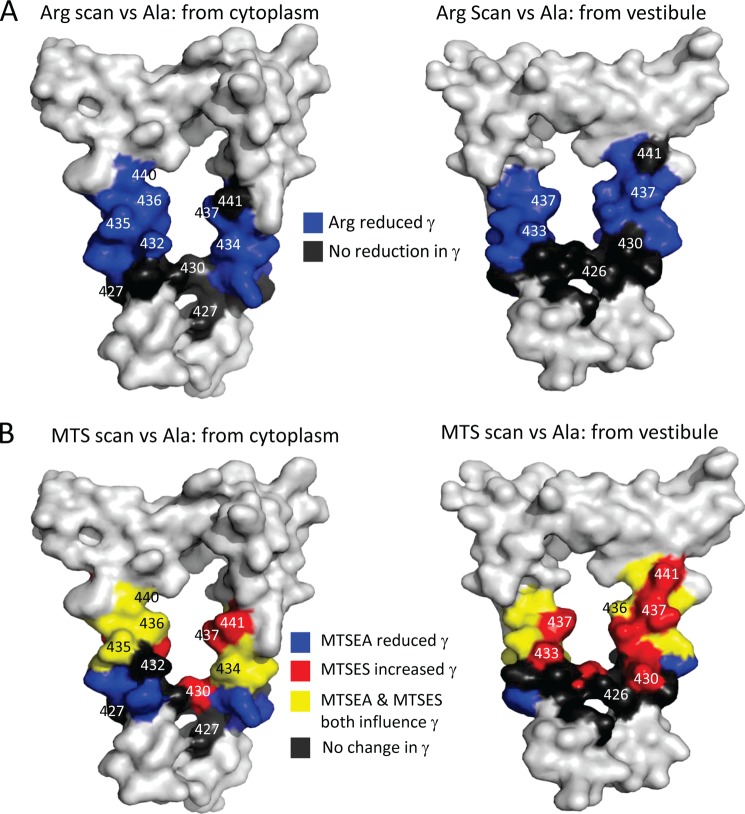
Structural models of Cys-loop receptors based on homology with the Torpedo marmorata nicotinic acetylcholine receptor infer the existence of cytoplasmic portals within the conduction pathway framed by helical amphipathic regions (termed membrane-associated (MA) helices) of adjacent intracellular M3-M4 loops. Consistent with these models, two arginine residues (Arg(436) and Arg(440)) within the MA helix of 5-hydroxytryptamine type 3A (5-HT3A) receptors act singularly as rate-limiting determinants of single-channel conductance (γ). However, there is little conservation in primary amino acid sequences across the cytoplasmic loops of Cys-loop receptors, limiting confidence in the fidelity of this particular aspect of the 5-HT3A receptor model. We probed the majority of residues within the MA helix of the human 5-HT3A subunit using alanine- and arginine-scanning mutagenesis and the substituted cysteine accessibility method to determine their relative influences upon γ. Numerous residues, prominently those at the 435, 436, 439, and 440 positions, were found to markedly influence γ. This approach yielded a functional map of the 5-HT3A receptor portals, which agrees well with the homology model.
Keywords: Cys-loop Receptors; Neurotransmitter Receptors; Nicotinic Acetylcholine Receptors; Patch Clamp Electrophysiology; Pentameric Ligand-gated Ion Channel; Serotonin.
Figures
References
-
- Millar N. S., Gotti C. (2009) Diversity of vertebrate nicotinic acetylcholine receptors. Neuropharmacology 56, 237–246 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
