

What pathogens have taught us about posttranslational modifications
- PMID: 24034613
- PMCID: PMC5785091
- DOI: 10.1016/j.chom.2013.07.008
What pathogens have taught us about posttranslational modifications
Abstract
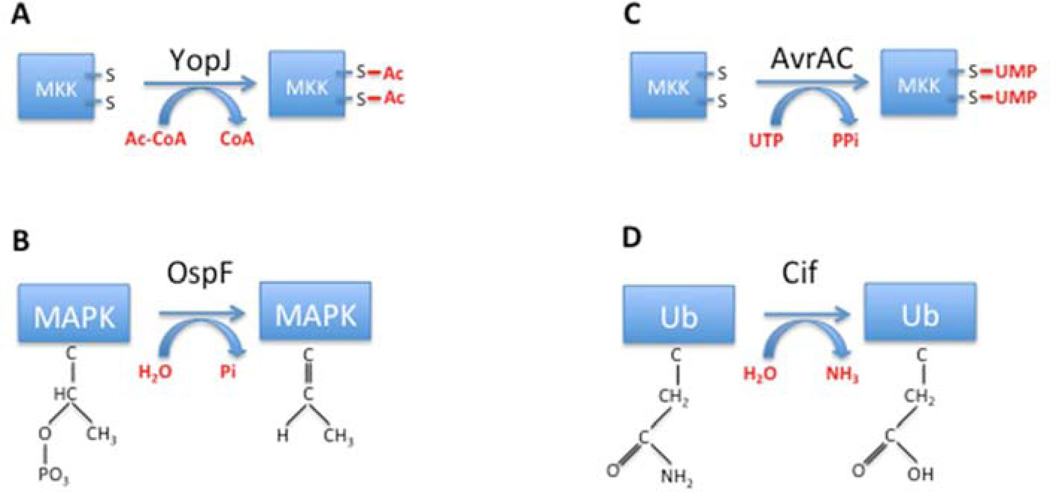
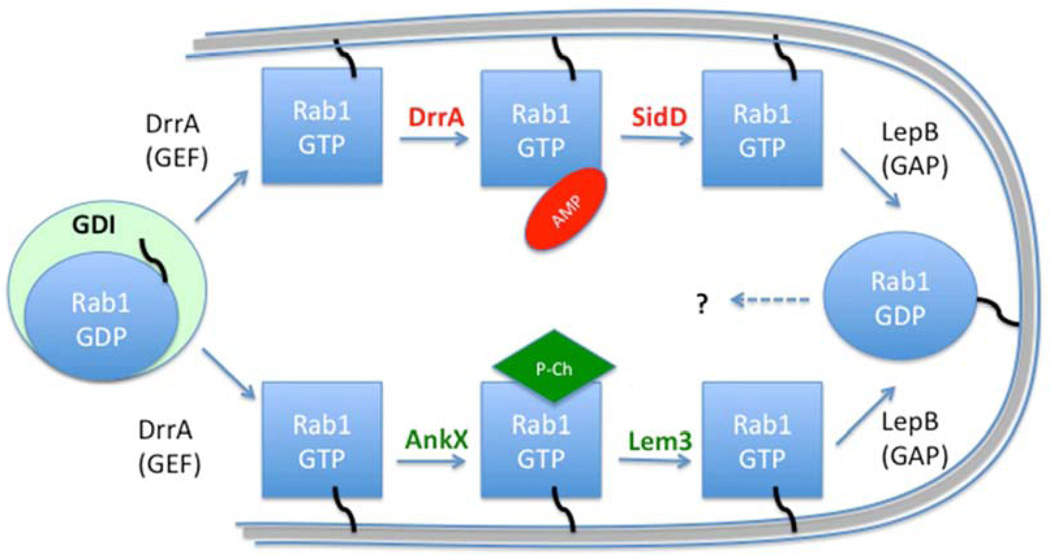
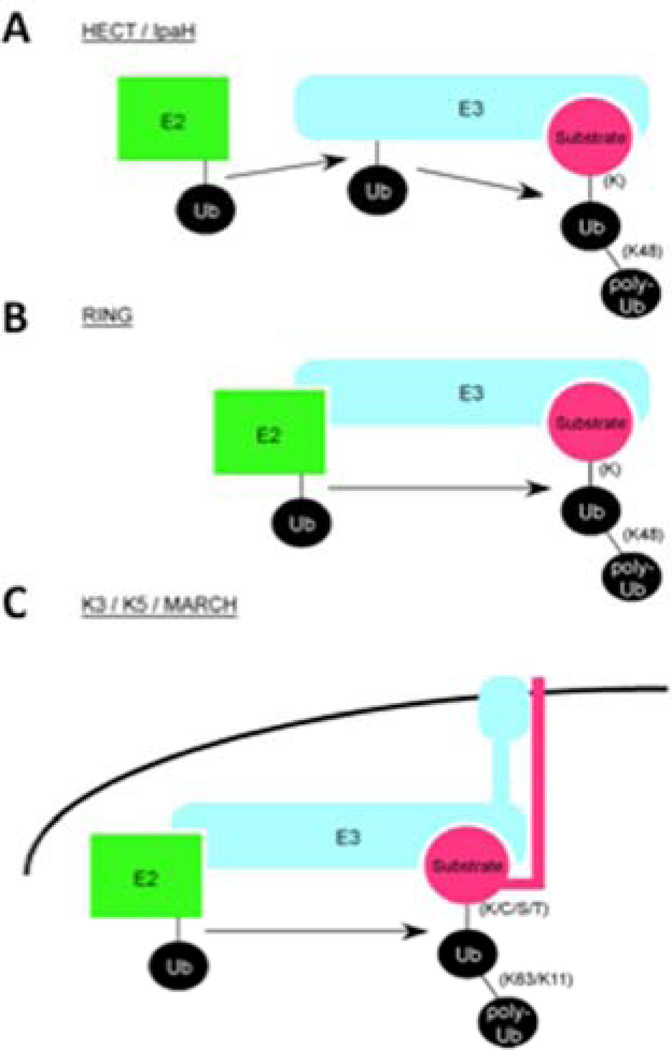
Pathogens use various mechanisms to manipulate host processes to promote infection. Decades of research on pathogens have revealed not only the molecular mechanisms that these microbes use to replicate and survive within host cells, but also seminal information on how host signaling machinery regulates cellular processes. Among these discoveries are mechanisms involving posttranslational modifications that alter the activity, localization, or interactions of the modified protein. Herein, we examine how pathogens have contributed to our basic understanding of three posttranslational modifications: phosphorylation, NMPylation, and ubiquitylation. Over the years, technologies, techniques and research tools have developed side by side with the study of pathogens, facilitating the discovery of protein modifications and furthering our understanding of how they contribute to both infection and cellular functions.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures
References
-
- Barford D. Protein phosphatases. Curr Opin Struct Biol. 1995;5:728–734. - PubMed
-
- Boggon TJ, Eck MJ. Structure and regulation of Src family kinases. Oncogene. 2004;23:7918–7927. - PubMed
-
- Bolin I, Wolf-Watz H. The plasmid-encoded Yop2b protein of Yersinia pseudotuberculosis is a virulence determinant regulated by calcium and temperature at the level of transcription. Mol Microbiol. 1988;2:237–245. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
