

Identifying Yersinia YopH-targeted signal transduction pathways that impair neutrophil responses during in vivo murine infection
- PMID: 24034616
- PMCID: PMC3789382
- DOI: 10.1016/j.chom.2013.08.013
Identifying Yersinia YopH-targeted signal transduction pathways that impair neutrophil responses during in vivo murine infection
Abstract
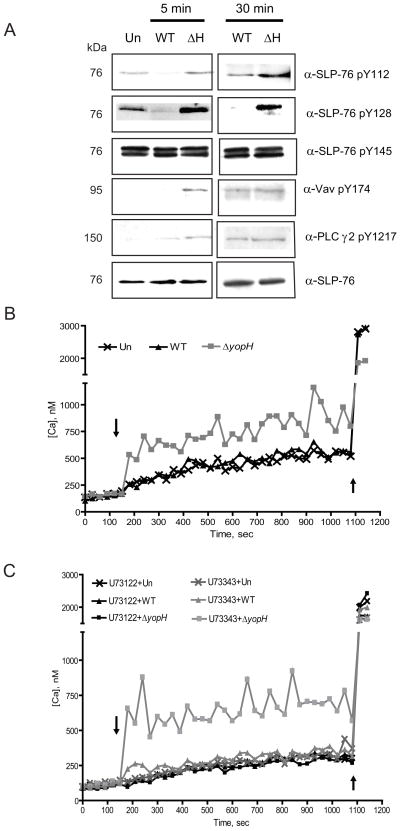
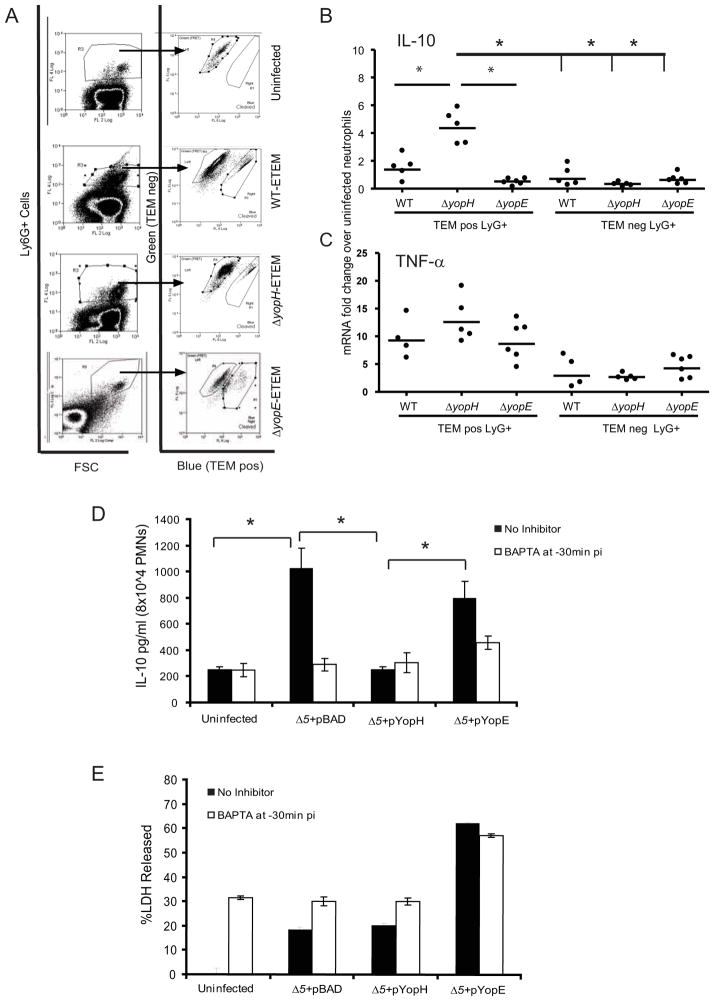
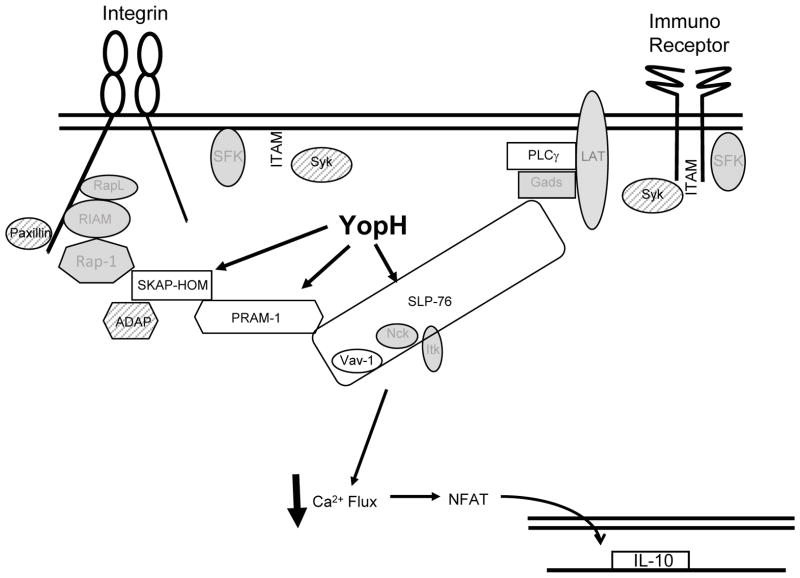
Identifying molecular targets of Yersinia virulence effectors, or Yops, during animal infection is challenging because few cells are targeted by Yops in an infected organ, and isolating these sparse effector-containing cells is difficult. YopH, a tyrosine phosphatase, is essential for full virulence of Yersinia. Investigating the YopH-targeted signal transduction pathway(s) in neutrophils during infection of a murine host, we find that several host proteins, including the essential signaling adaptor SLP-76, are dephosphorylated in the presence of YopH in neutrophils isolated from infected tissues. YopH inactivated PRAM-1/SKAP-HOM and the SLP-76/Vav/PLCγ2 signal transduction axes, leading to an inhibition of calcium response in isolated neutrophils. Consistent with a failure to mount a calcium response, IL-10 production was reduced in neutrophils containing YopH from infected tissues. Finally, a yopH mutant survived better in the absence of neutrophils, indicating that neutrophil inactivation by YopH by targeting PRAM-1/SKAP-HOM and SLP-76/Vav/PLCγ2 signaling hubs may be critical for Yersinia survival.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures
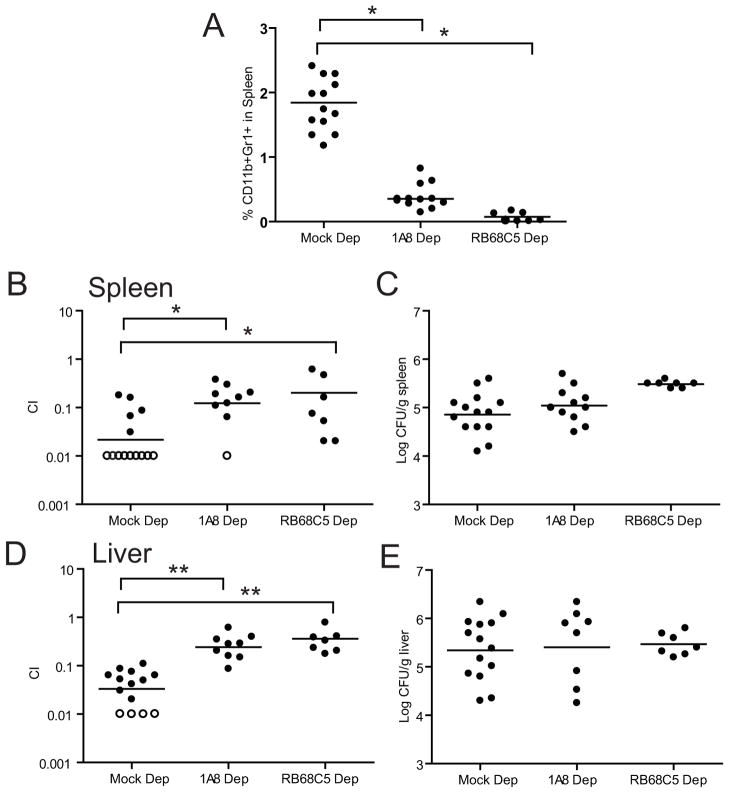
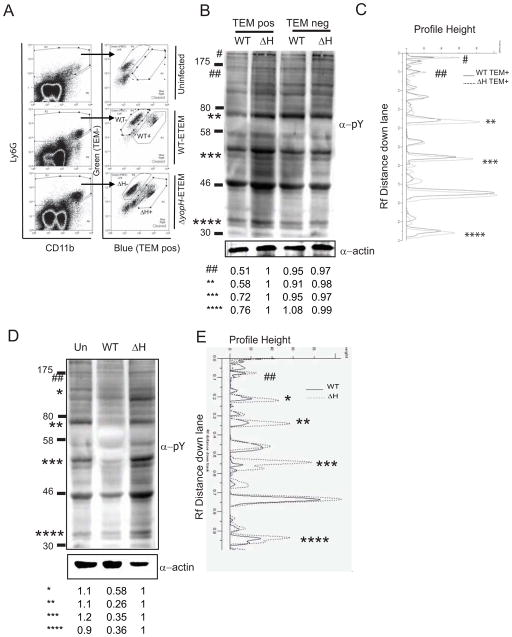
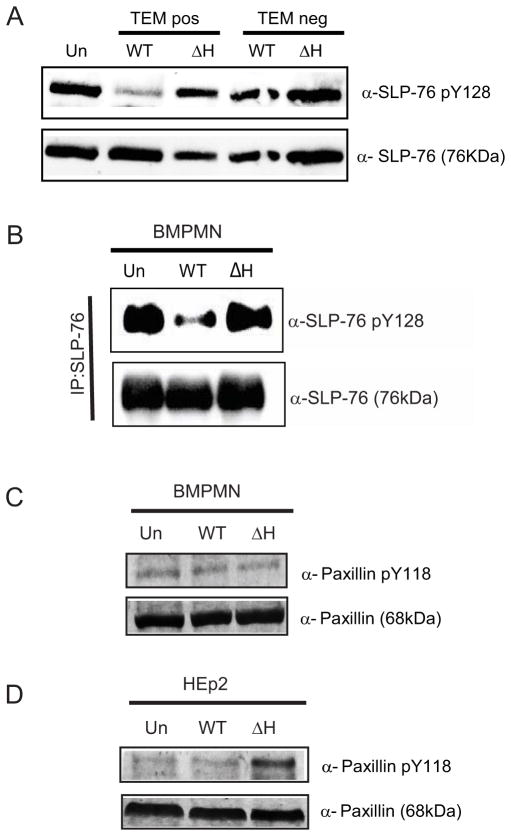
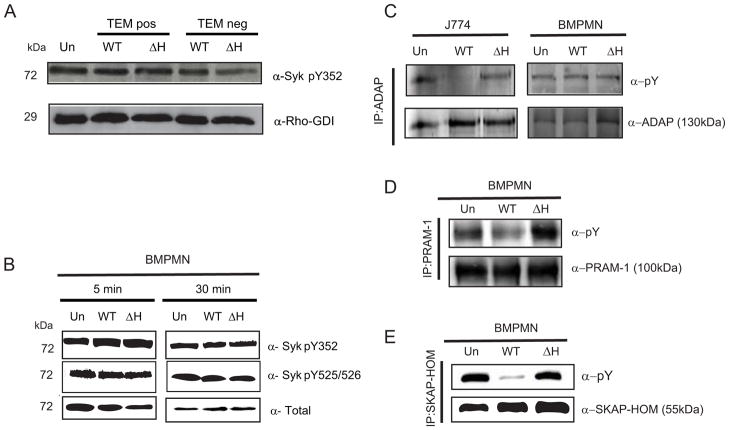
References
-
- Alonso A, Bottini N, Bruckner S, Rahmouni S, Williams S, Schoenberger SP, Mustelin T. Lck dephosphorylation at Tyr-394 and inhibition of T cell antigen receptor signaling by Yersinia phosphatase YopH. J Biol Chem. 2004;279:4922–4928. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
