

Efficient disruption of Zebrafish genes using a Gal4-containing gene trap
- PMID: 24034702
- PMCID: PMC3848861
- DOI: 10.1186/1471-2164-14-619
Efficient disruption of Zebrafish genes using a Gal4-containing gene trap
Abstract
Background: External development and optical transparency of embryos make zebrafish exceptionally suitable for in vivo insertional mutagenesis using fluorescent proteins to visualize expression patterns of mutated genes. Recently developed Gene Breaking Transposon (GBT) vectors greatly improve the fidelity and mutagenicity of transposon-based gene trap vectors.
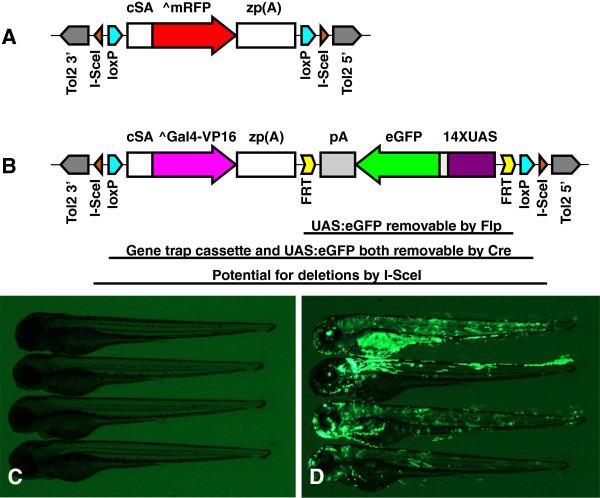
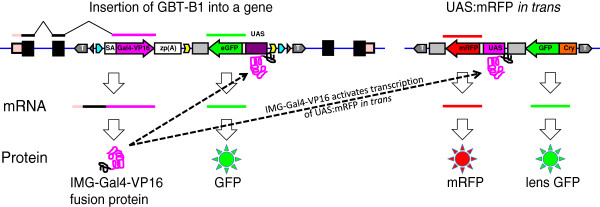
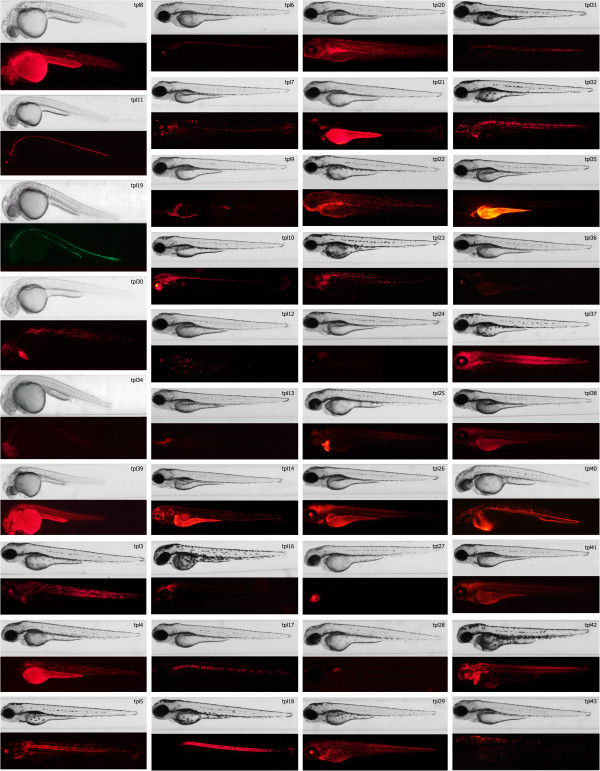
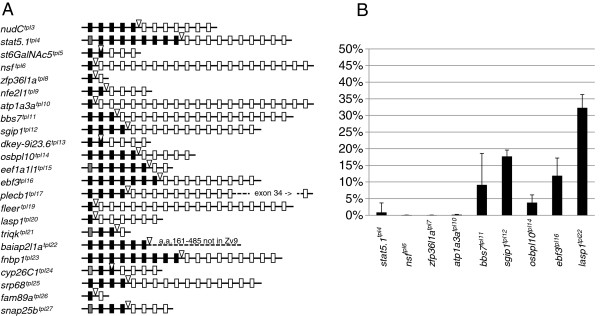
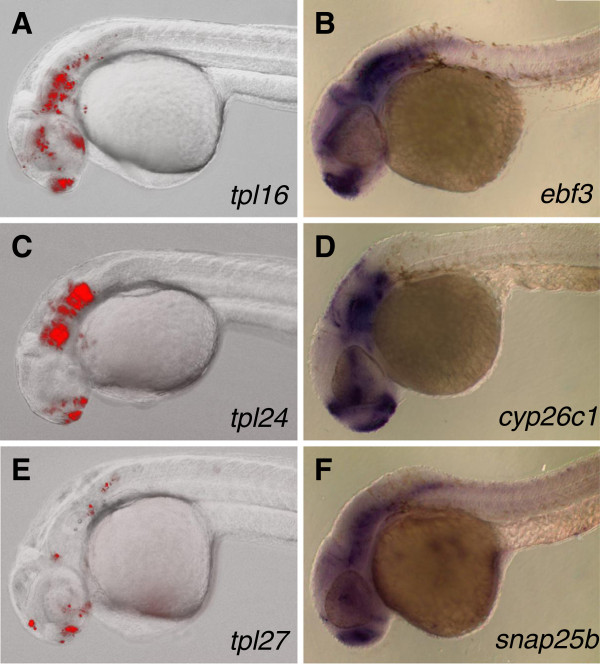
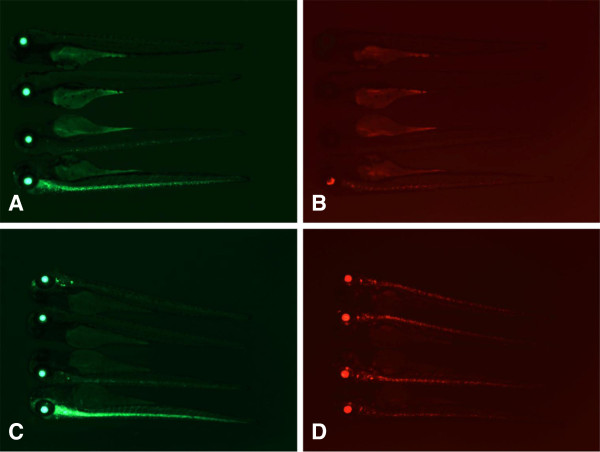
Results: We constructed and tested a bipartite GBT vector with Gal4-VP16 as the primary gene trap reporter. Our vector also contains a UAS:eGFP cassette for direct detection of gene trap events by fluorescence. To confirm gene trap events, we generated a UAS:mRFP tester line. We screened 270 potential founders and established 41 gene trap lines. Three of our gene trap alleles display homozygous lethal phenotypes ranging from embryonic to late larval: nsf( tpl6), atp1a3a(tpl10) and flr(tpl19). Our gene trap cassette is flanked by direct loxP sites, which enabled us to successfully revert nsf( tpl6), atp1a3a(tpl10) and flr(tpl19) gene trap alleles by injection of Cre mRNA. The UAS:eGFP cassette is flanked by direct FRT sites. It can be readily removed by injection of Flp mRNA for use of our gene trap alleles with other tissue-specific GFP-marked lines. The Gal4-VP16 component of our vector provides two important advantages over other GBT vectors. The first is increased sensitivity, which enabled us to detect previously unnoticed expression of nsf in the pancreas. The second advantage is that all our gene trap lines, including integrations into non-essential genes, can be used as highly specific Gal4 drivers for expression of other transgenes under the control of Gal4 UAS.
Conclusions: The Gal4-containing bipartite Gene Breaking Transposon vector presented here retains high specificity for integrations into genes, high mutagenicity and revertibility by Cre. These features, together with utility as highly specific Gal4 drivers, make gene trap mutants presented here especially useful to the research community.
Figures
References
-
- Haffter P, Granato M, Brand M, Mullins MC, Hammerschmidt M, Kane DA, Odenthal J, van Eeden FJ, Jiang YJ, Heisenberg CP. et al. The identification of genes with unique and essential functions in the development of the zebrafish, Danio rerio. Development. 1996;123:1–36. - PubMed
-
- Driever W, Solnica-Krezel L, Schier AF, Neuhauss SC, Malicki J, Stemple DL, Stainier DY, Zwartkruis F, Abdelilah S, Rangini Z. et al. A genetic screen for mutations affecting embryogenesis in zebrafish. Development. 1996;123:37–46. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
