

Domain exchange at the 3' end of the gene encoding the fratricide meningococcal two-partner secretion protein A
- PMID: 24034852
- PMCID: PMC3848433
- DOI: 10.1186/1471-2164-14-622
Domain exchange at the 3' end of the gene encoding the fratricide meningococcal two-partner secretion protein A
Abstract
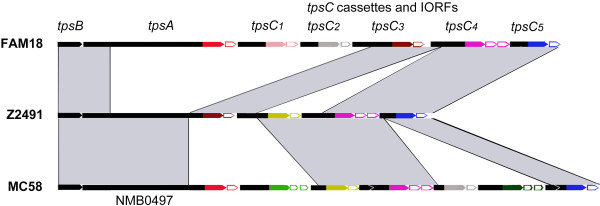
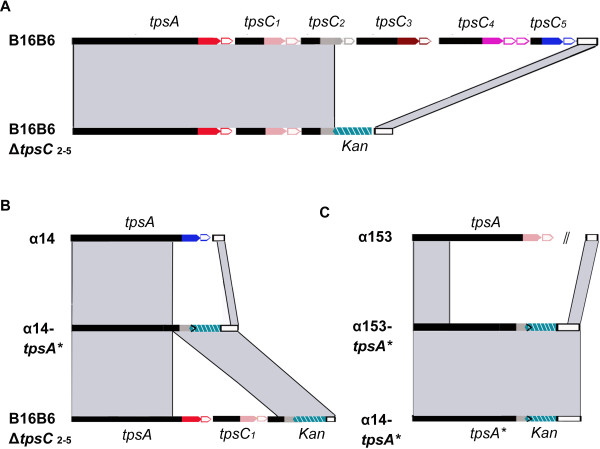
Background: Two-partner secretion systems in Gram-negative bacteria consist of an outer membrane protein TpsB that mediates the secretion of a cognate TpsA protein into the extracellular milieu. TpsA proteins have diverse, often virulence-related functions, and some of them inhibit the growth of related bacteria. In Neisseria meningitidis, several functions have been attributed to the TpsA proteins. Downstream of the tpsB and tpsA genes, several shorter tpsA-related gene cassettes, called tpsC, are located interspersed with intervening open-reading frames (IORFs). It has been suggested that the tpsC cassettes may recombine with the tpsA gene as a mechanism of antigenic variation. Here, we investigated (i) whether TpsA of N. meningitidis also has growth-inhibitory properties, (ii) whether tpsC cassettes recombine with the tpsA gene, and (iii) what the consequences of such recombination events might be.
Results: We demonstrate that meningococcal TpsA has growth-inhibitory properties and that the IORF located immediately downstream of tpsA confers immunity to the producing strain. Although bioinformatics analysis suggests that recombination between tpsC cassettes and tpsA occurs, detailed analysis of the tpsA gene in a large collection of disease isolates of three clonal complexes revealed that the frequency is very low and cannot be a mechanism of antigenic variation. However, recombination affected growth inhibition. In vitro experiments revealed that recombination can be mediated through acquirement of tpsC cassettes from the environment and it identified the regions involved in the recombination.
Conclusions: Meningococcal TpsA has growth-inhibitory properties. Recombination between tpsA and tpsC cassettes occurs in vivo but is rare and has consequences for growth inhibition. A recombination model is proposed and we propose that the main goal of recombination is the collection of new IORFs for protection against a variety of TpsA proteins.
Figures



Similar articles
-
Fratricide activity of MafB protein of N. meningitidis strain B16B6.BMC Microbiol. 2015 Aug 5;15:156. doi: 10.1186/s12866-015-0493-6. BMC Microbiol. 2015. PMID: 26242409 Free PMC article.
-
Two-partner secretion systems of Neisseria meningitidis associated with invasive clonal complexes.Infect Immun. 2008 Oct;76(10):4649-58. doi: 10.1128/IAI.00393-08. Epub 2008 Aug 4. Infect Immun. 2008. PMID: 18678657 Free PMC article.
-
System specificity of the TpsB transporters of coexpressed two-partner secretion systems of Neisseria meningitidis.J Bacteriol. 2013 Feb;195(4):788-97. doi: 10.1128/JB.01355-12. Epub 2012 Dec 7. J Bacteriol. 2013. PMID: 23222722 Free PMC article.
-
Two-Partner Secretion: Combining Efficiency and Simplicity in the Secretion of Large Proteins for Bacteria-Host and Bacteria-Bacteria Interactions.Front Cell Infect Microbiol. 2017 May 9;7:148. doi: 10.3389/fcimb.2017.00148. eCollection 2017. Front Cell Infect Microbiol. 2017. PMID: 28536673 Free PMC article. Review.
-
Microevolution and epidemic spread of serogroup A Neisseria meningitidis--a review.Gene. 1997 Jun 11;192(1):135-40. doi: 10.1016/s0378-1119(97)00083-8. Gene. 1997. PMID: 9224883 Review.
Cited by
-
Editorial: Pathogenic Neisseria: Pathogenicity, vaccines, and antibiotic resistance.Front Cell Infect Microbiol. 2023 Jan 5;12:1119244. doi: 10.3389/fcimb.2022.1119244. eCollection 2022. Front Cell Infect Microbiol. 2023. PMID: 36683679 Free PMC article. No abstract available.
-
Meningococcal Two-Partner Secretion Systems and Their Association with Outcome in Patients with Meningitis.Infect Immun. 2016 Aug 19;84(9):2534-40. doi: 10.1128/IAI.00160-16. Print 2016 Sep. Infect Immun. 2016. PMID: 27324486 Free PMC article.
-
Molecular interactions between Neisseria meningitidis and its human host.Cell Microbiol. 2019 Nov;21(11):e13063. doi: 10.1111/cmi.13063. Epub 2019 Jun 13. Cell Microbiol. 2019. PMID: 31167044 Free PMC article. Review.
-
HrpA anchors meningococci to the dynein motor and affects the balance between apoptosis and pyroptosis.J Biomed Sci. 2022 Jun 28;29(1):45. doi: 10.1186/s12929-022-00829-8. J Biomed Sci. 2022. PMID: 35765029 Free PMC article.
-
The structure of a contact-dependent growth-inhibition (CDI) immunity protein from Neisseria meningitidis MC58.Acta Crystallogr F Struct Biol Commun. 2015 Jun;71(Pt 6):702-9. doi: 10.1107/S2053230X15006585. Epub 2015 May 20. Acta Crystallogr F Struct Biol Commun. 2015. PMID: 26057799 Free PMC article.
References
-
- Bentley SD, Vernikos GS, Snyder LAS, Churcher C, Arrowsmith C, Chillingworth T, Cronin A, Davis PH, Holroyd NE, Jagels K, Maddison M, Moule S, Rabbinowitsch E, Sharp S, Unwin L, Whitehead S, Quail MA, Achtman M, Barrell B, Saunders NJ, Parkhill J. Meningococcal genetic variation mechanisms viewed through comparative analysis of serogroup C strain FAM18. PLoS Genet. 2007;3:e23. doi: 10.1371/journal.pgen.0030023. - DOI - PMC - PubMed
-
- Santoyo G, Romero D. Gene conversion and concerted evolution in bacterial genomes. FEMS Microbiol Rev. 2005;29:169–183. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
