

Synapse maturation by activity-dependent ectodomain shedding of SIRPα
- PMID: 24036914
- PMCID: PMC3820962
- DOI: 10.1038/nn.3516
Synapse maturation by activity-dependent ectodomain shedding of SIRPα
Abstract
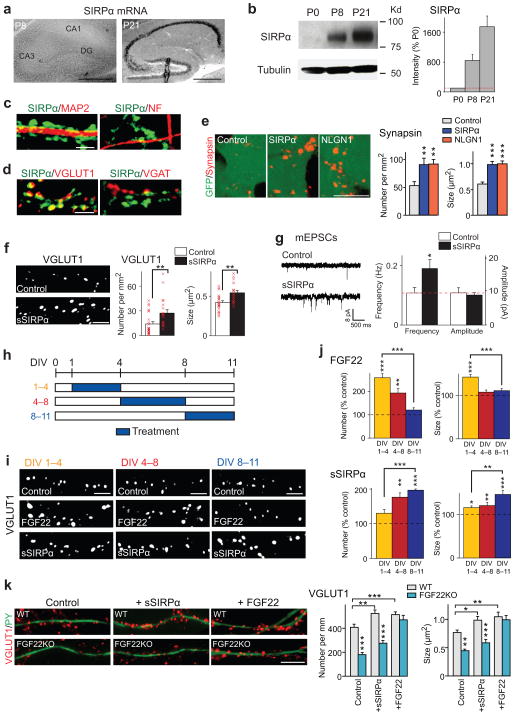
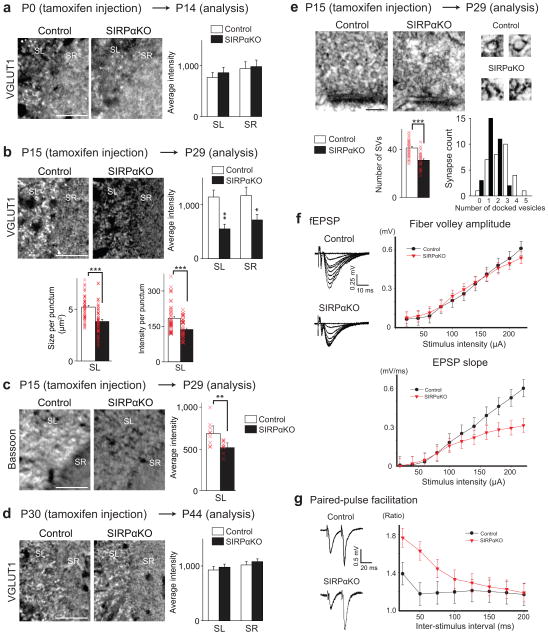
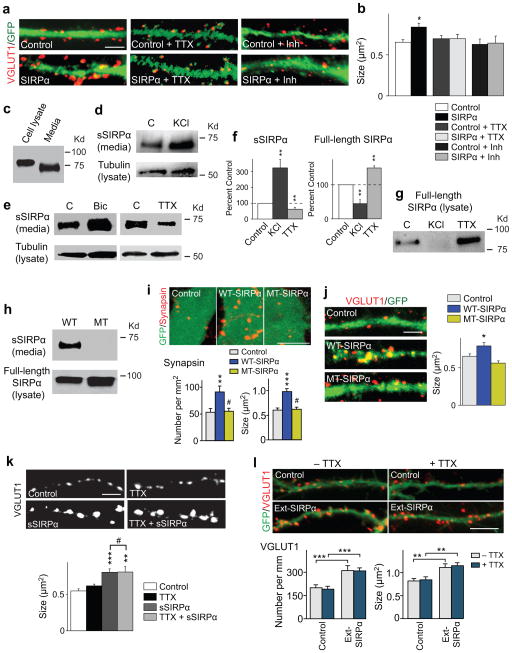
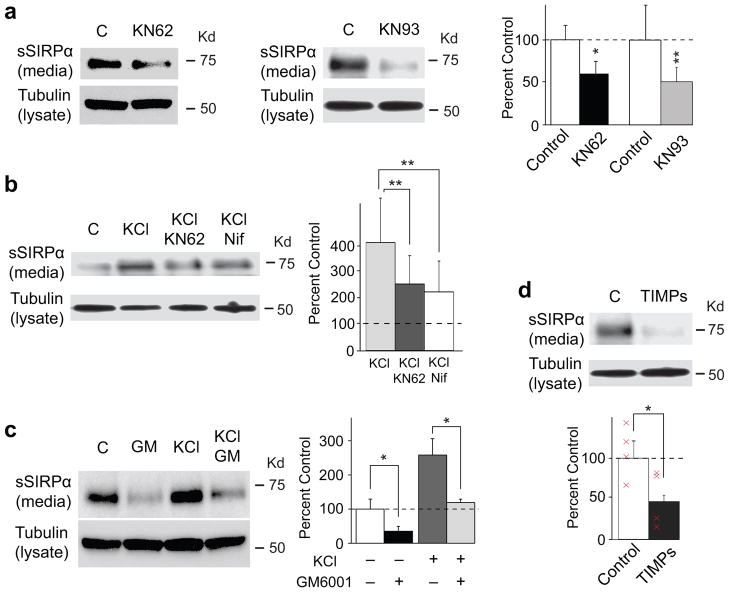
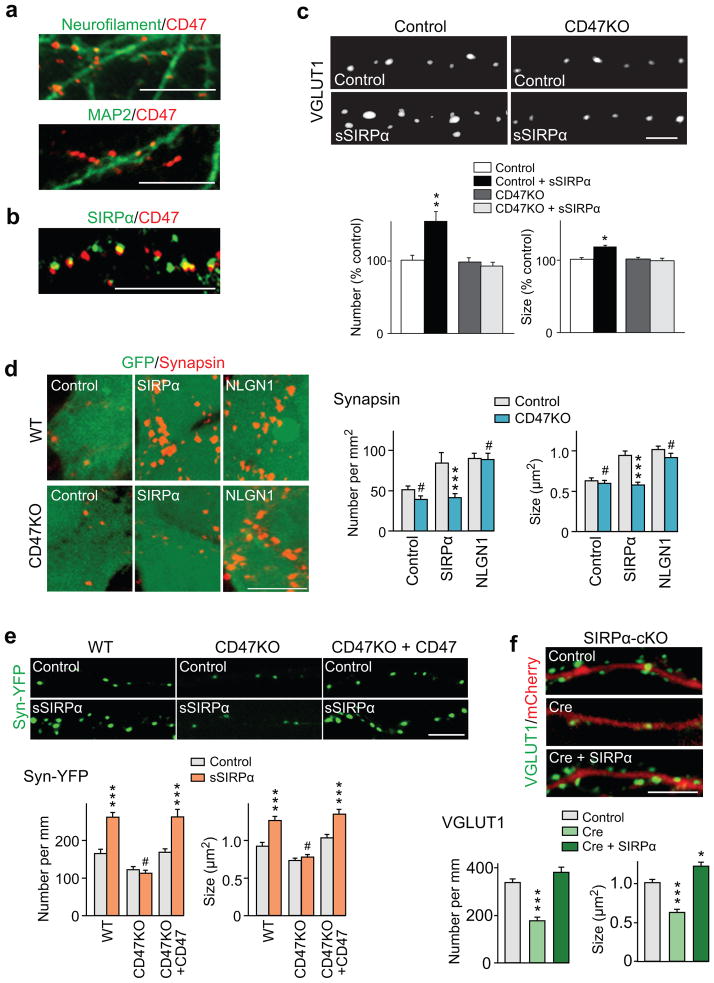
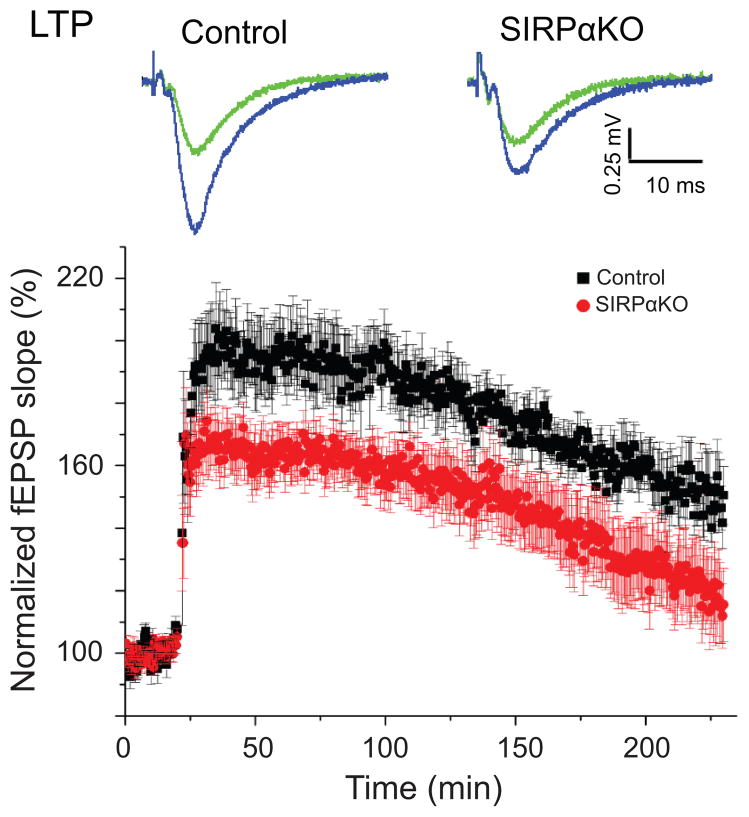
Formation of appropriate synaptic connections is critical for proper functioning of the brain. After initial synaptic differentiation, active synapses are stabilized by neural activity-dependent signals to establish functional synaptic connections. However, the molecular mechanisms underlying activity-dependent synapse maturation remain to be elucidated. Here we show that activity-dependent ectodomain shedding of signal regulatory protein-α (SIRPα) mediates presynaptic maturation. Two target-derived molecules, fibroblast growth factor 22 and SIRPα, sequentially organize the glutamatergic presynaptic terminals during the initial synaptic differentiation and synapse maturation stages, respectively, in the mouse hippocampus. SIRPα drives presynaptic maturation in an activity-dependent fashion. Remarkably, neural activity cleaves the extracellular domain of SIRPα, and the shed ectodomain in turn promotes the maturation of the presynaptic terminal. This process involves calcium/calmodulin-dependent protein kinase, matrix metalloproteinases and the presynaptic receptor CD47. Finally, SIRPα-dependent synapse maturation has an impact on synaptic function and plasticity. Thus, ectodomain shedding of SIRPα is an activity-dependent trans-synaptic mechanism for the maturation of functional synapses.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
Comment in
-
Synaptic uSIRPation: the active neuron reigns over presynaptic partners.Nat Neurosci. 2013 Oct;16(10):1361-2. doi: 10.1038/nn.3523. Nat Neurosci. 2013. PMID: 24067286 No abstract available.
References
-
- Kasai H, Fukuda M, Watanabe S, Hayashi-Takagi A, Noguchi J. Structural dynamics of dendritic spines in memory and cognition. Trends Neurosci. 2010;33:121–129. - PubMed
-
- Sanes JR, Lichtman JW. Development of the vertebrate neuromuscular junction. Annu Rev Neurosci. 1999;22:389–442. - PubMed
-
- Waites CL, Craig AM, Garner CC. Mechanisms of vertebrate synaptogenesis. Annu Rev Neurosci. 2005;28:251–274. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
