

Comparative secretome analysis of epithelial and mesenchymal subpopulations of head and neck squamous cell carcinoma identifies S100A4 as a potential therapeutic target
- PMID: 24037664
- PMCID: PMC3861723
- DOI: 10.1074/mcp.M113.029587
Comparative secretome analysis of epithelial and mesenchymal subpopulations of head and neck squamous cell carcinoma identifies S100A4 as a potential therapeutic target
Abstract
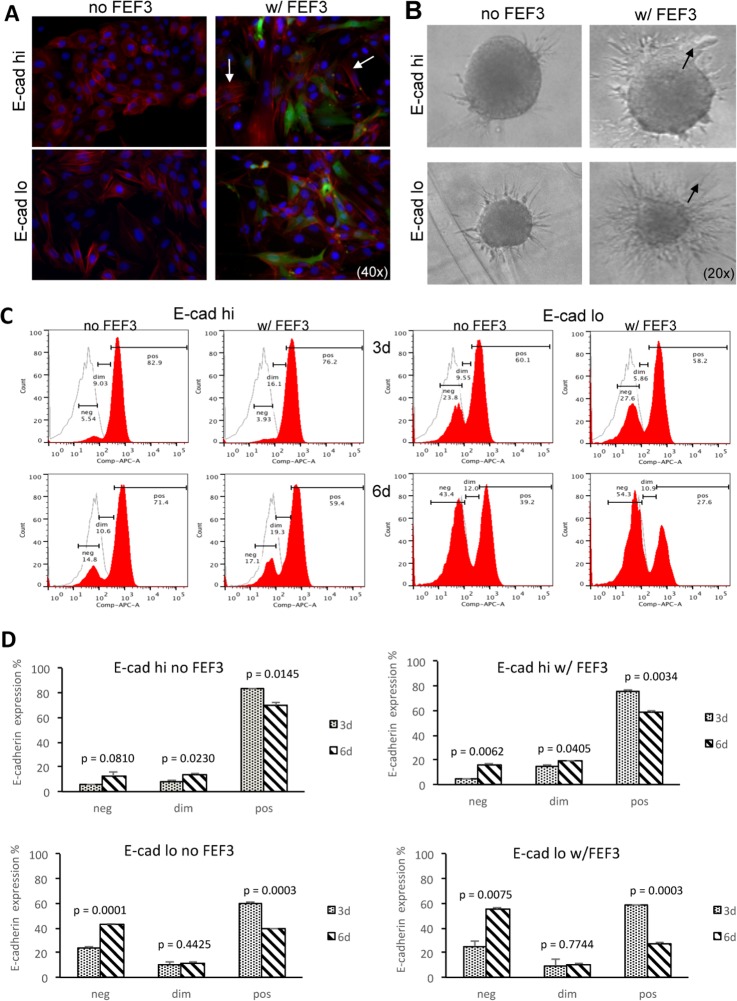
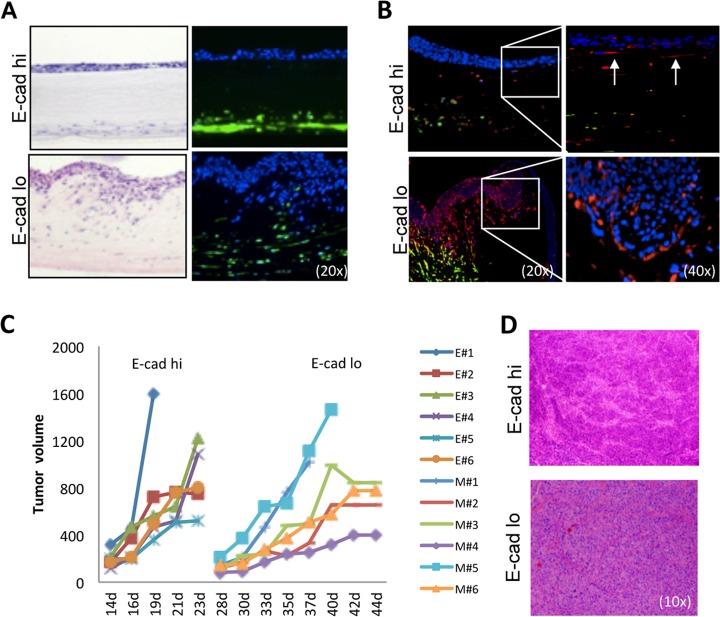
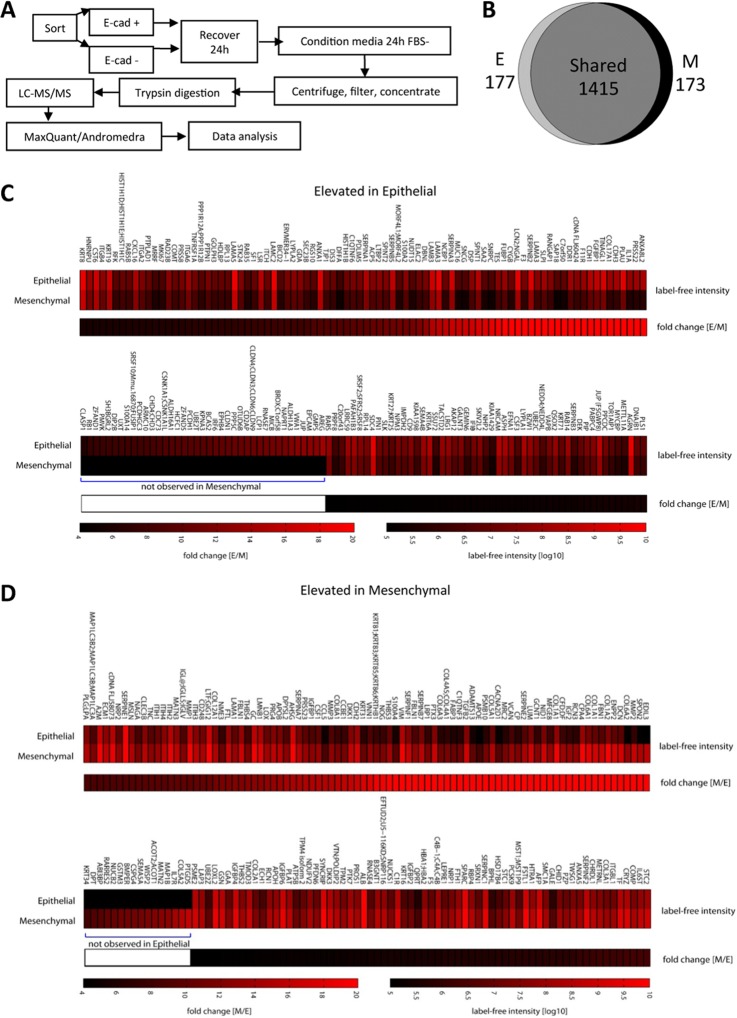
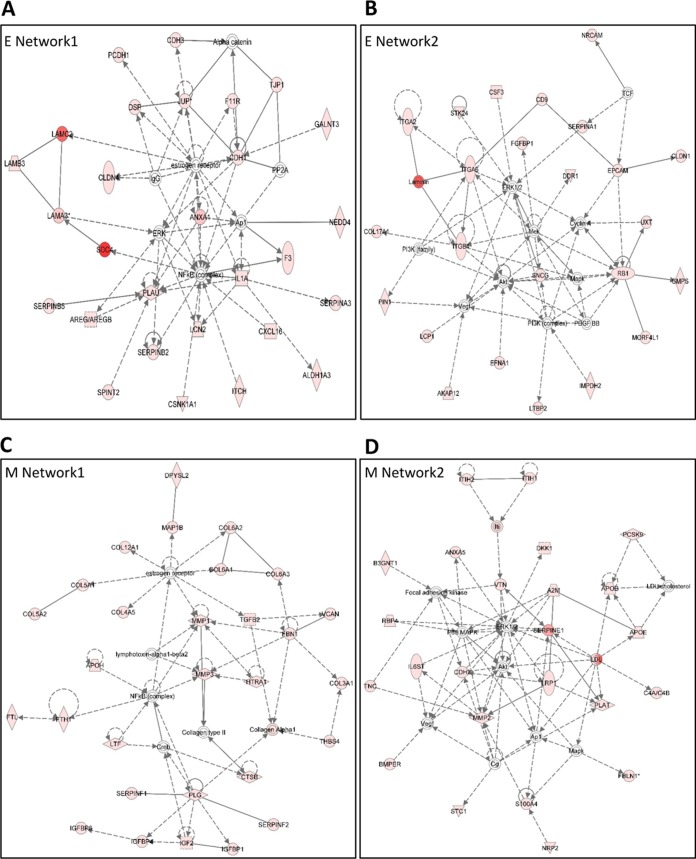
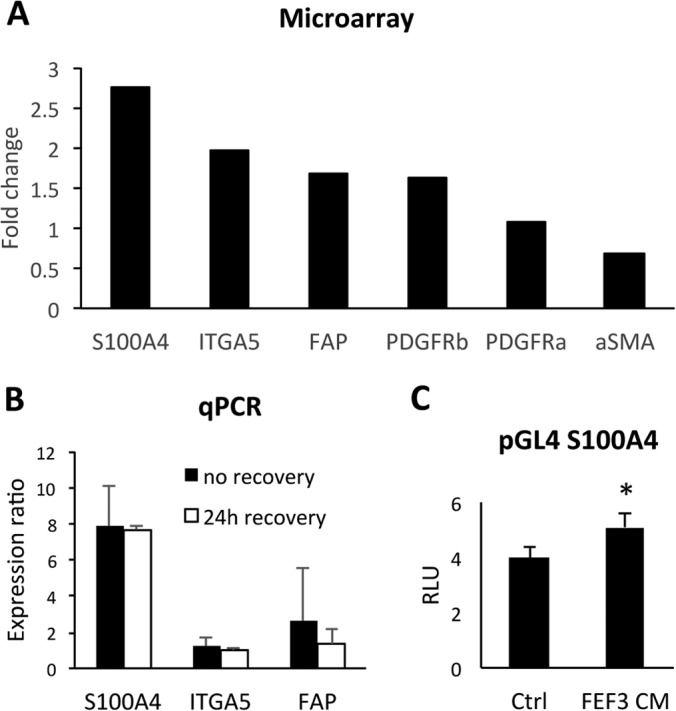
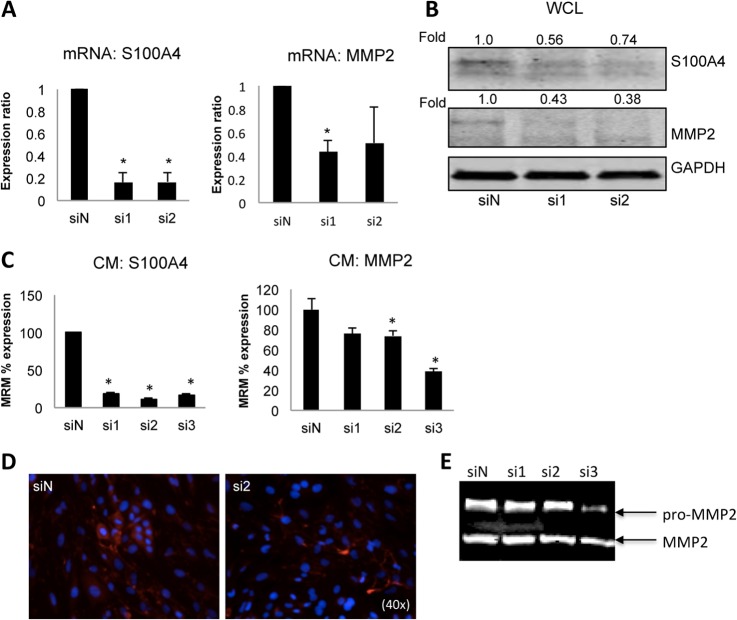
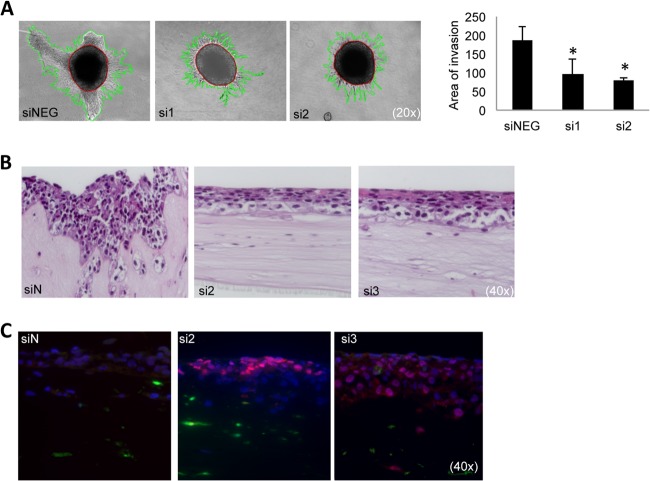
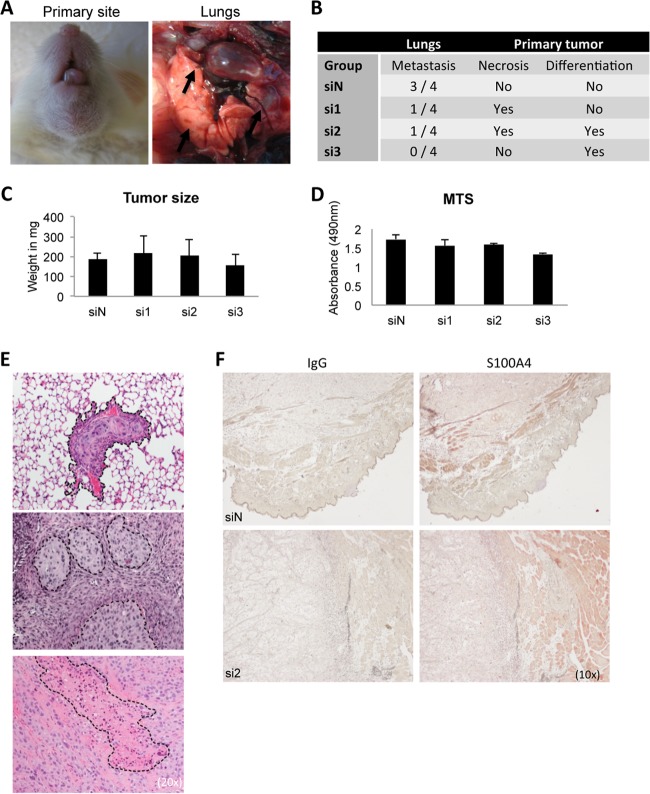
Epithelial-mesenchymal transition (EMT) is a key contributor in tumor progression and metastasis. EMT produces cellular heterogeneity within head and neck squamous cell carcinomas (HNSCC) by creating a phenotypically distinct mesenchymal subpopulation that is resistant to conventional therapies. In this study, we systematically characterized differences in the secretomes of E-cadherin high epithelial-like and E-cadherin low mesenchymal-like subpopulations using unbiased and targeted proteomics. A total 1765 proteins showed significant changes with 177 elevated in the epithelial subpopulation and 173 elevated in the mesenchymal cells. Key nodes in affected networks included NFκB, Akt, and ERK, and most implicated cellular components involved various aspects of the extracellular matrix. In particular, large changes were observed in multiple collagens with most affected collagens at much higher abundance levels in the mesenchymal subpopulation. These cells also exhibited a secretome profile resembling that of cancer-associated fibroblastic cells (CAF). S100A4, a commonly used marker for cancer-associated fibroblastic cells, was elevated more than 20-fold in the mesenchymal cells and this increase was further verified at the transcriptome level. S100A4 is a known mediator of EMT, leading to metastasis and EMT has been proposed as a potential source of cancer-associated fibroblastic cells in solid tumors. S100A4 knockdown by small interfering RNA led to decreased expression, secretion and activity of matrix metalloproteinase 2, as verified by quantitative PCR, multiple reaction monitoring and zymography analyses, and reduced invasion in collagen-embedded spheroids. Further confirmation in three-dimensional organotypic reconstructs showed less invasion and advanced differentiation in the S100A4 RNA interference samples. Orthotopic metastasis model, developed to validate the findings in vivo, demonstrated a decrease in spontaneous metastasis and augmented differentiation in the primary tumor in siS100A4 xenografts. These results demonstrate the value of secretome profiling to evaluate phenotypic conversion and identify potential novel therapeutic targets such as S100A4.
Conflict of interest statement
Conflict of interest: The authors declare no conflict of interest.
Figures
References
-
- Basu D., Nguyen T. T., Montone K. T., Zhang G., Wang L. P., Diehl J. A., Rustgi A. K., Lee J. T., Weinstein G. S., Herlyn M. (2010) Evidence for mesenchymal-like sub-populations within squamous cell carcinomas possessing chemoresistance and phenotypic plasticity. Oncogene 29, 4170–4182 - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
