

Reciprocal regulation of microRNA-122 and c-Myc in hepatocellular cancer: role of E2F1 and transcription factor dimerization partner 2
- PMID: 24038073
- PMCID: PMC4199239
- DOI: 10.1002/hep.26712
Reciprocal regulation of microRNA-122 and c-Myc in hepatocellular cancer: role of E2F1 and transcription factor dimerization partner 2
Abstract
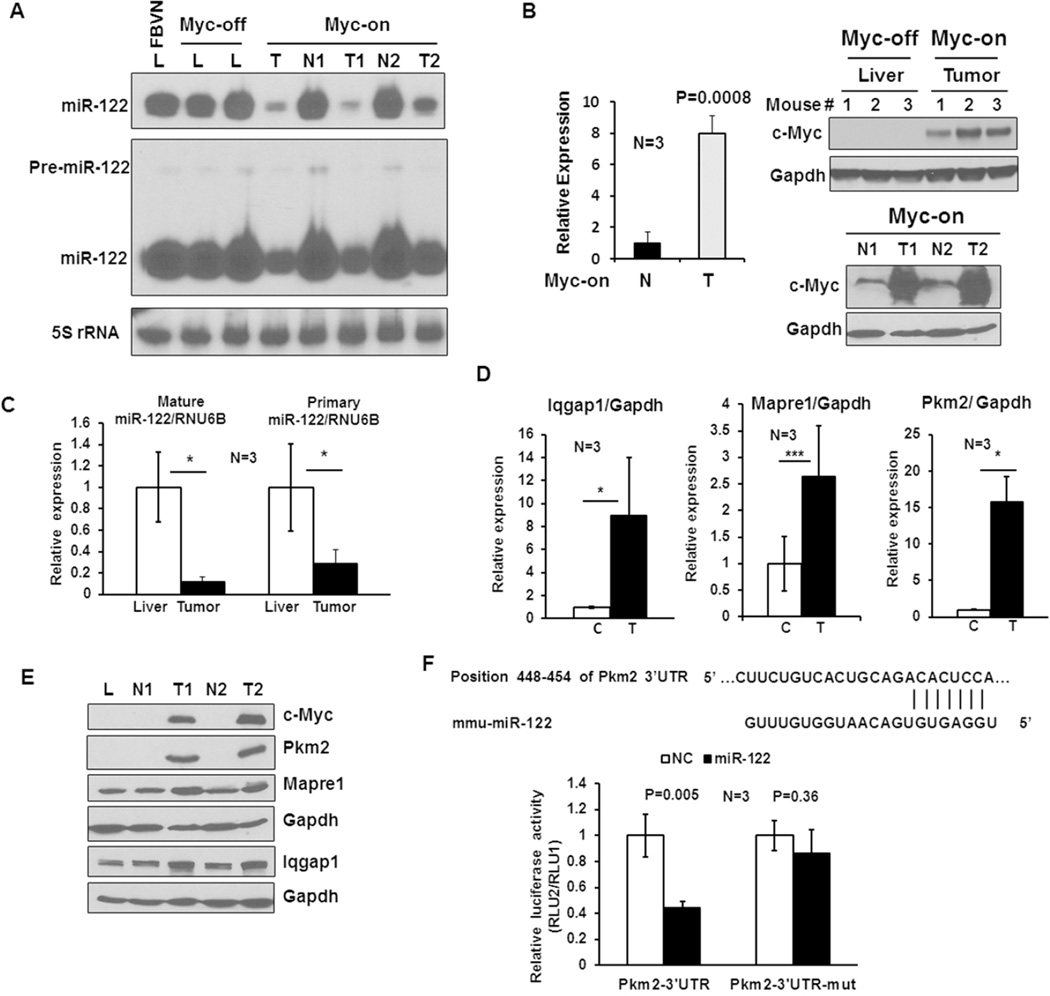
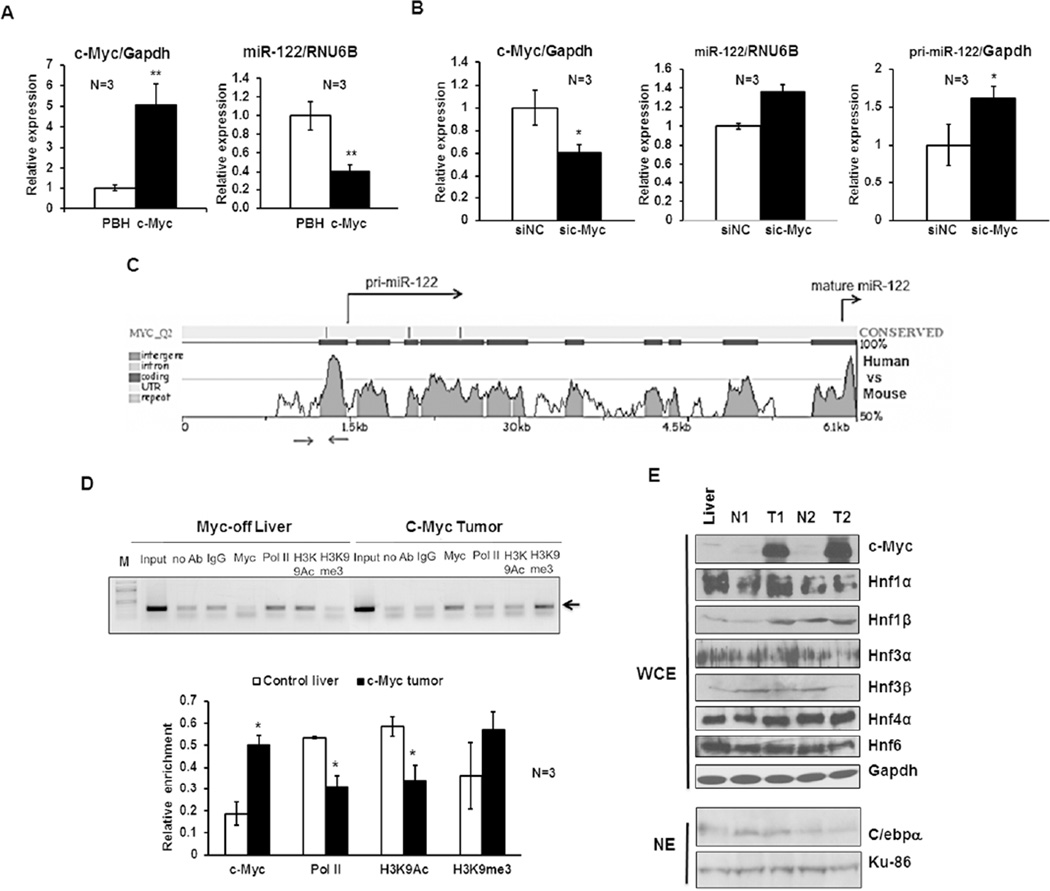
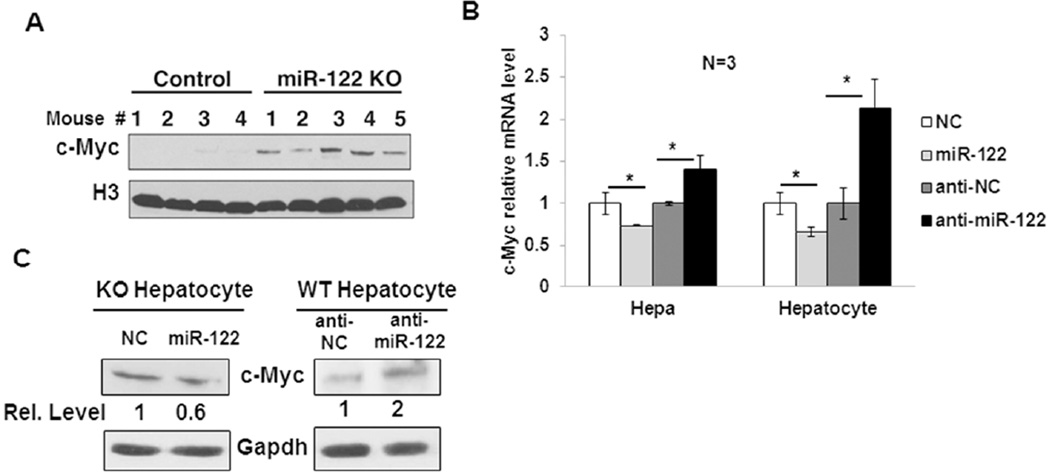
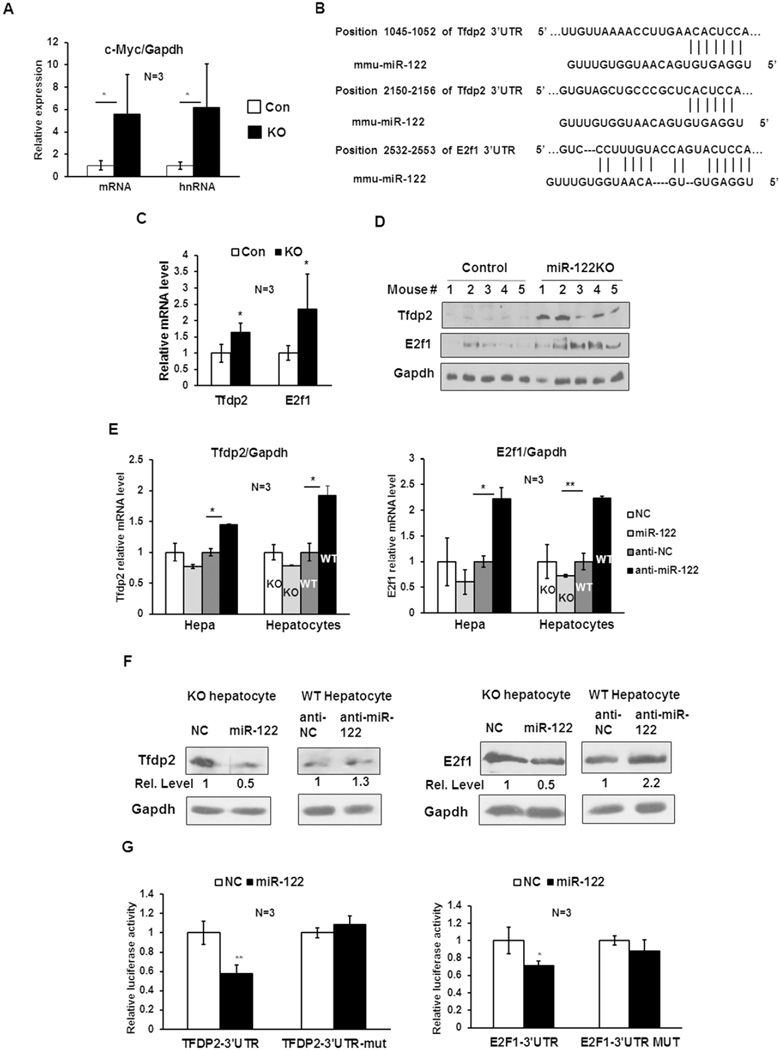
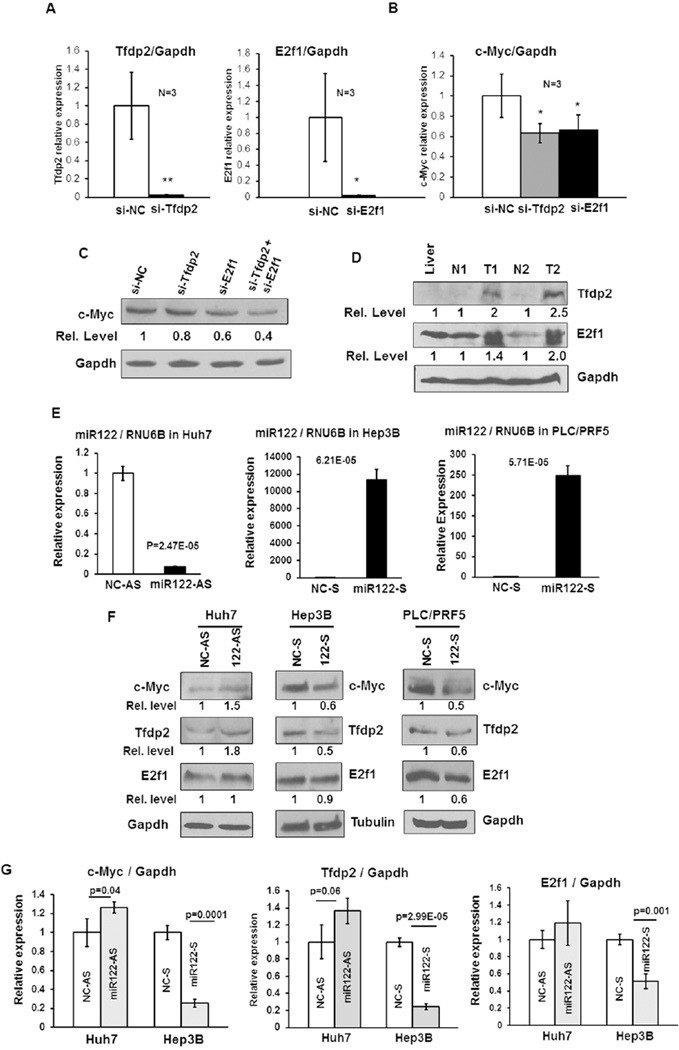
c-Myc is a well-known oncogene frequently up-regulated in different malignancies, whereas liver-specific microRNA (miR)-122, a bona fide tumor suppressor, is down-regulated in hepatocellular cancer (HCC). Here we explored the underlying mechanism of reciprocal regulation of these two genes. Real-time reverse-transcription polymerase chain reaction (RT-PCR) and northern blot analysis demonstrated reduced expression of the primary, precursor, and mature miR-122 in c-MYC-induced HCCs compared to the benign livers, indicating transcriptional suppression of miR-122 upon MYC overexpression. Indeed, chromatin immunoprecipitation (ChIP) assay showed significantly reduced association of RNA polymerase II and histone H3K9Ac, markers of active chromatin, with the miR-122 promoter in tumors relative to the c-MYC-uninduced livers, indicating transcriptional repression of miR-122 in c-MYC-overexpressing tumors. The ChIP assay also demonstrated a significant increase in c-Myc association with the miR-122 promoter region that harbors a conserved noncanonical c-Myc binding site in tumors compared to the livers. Ectopic expression and knockdown studies showed that c-Myc indeed suppresses expression of primary and mature miR-122 in hepatic cells. Additionally, Hnf-3β, a liver enriched transcription factor that activates miR-122 gene, was suppressed in c-MYC-induced tumors. Notably, miR-122 also repressed c-Myc transcription by targeting transcriptional activator E2f1 and coactivator Tfdp2, as evident from ectopic expression and knockdown studies and luciferase reporter assays in mouse and human hepatic cells.
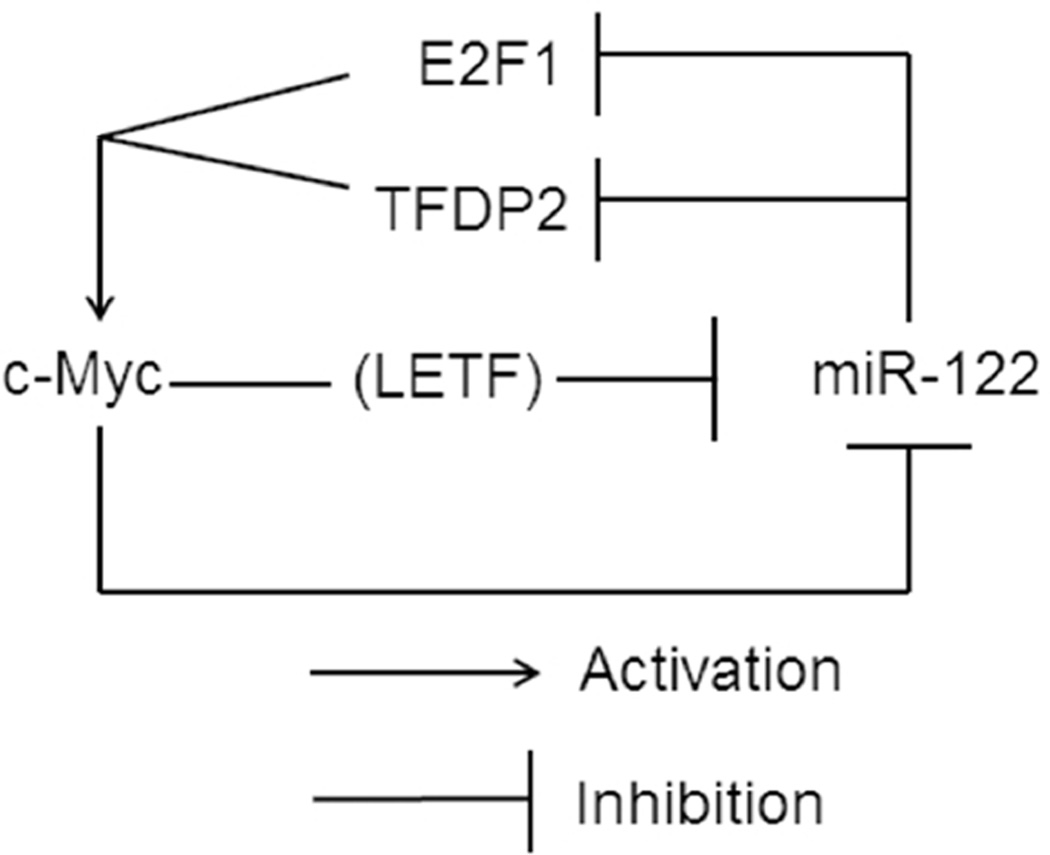
Conclusion: c-Myc represses miR-122 gene expression by associating with its promoter and by down-regulating Hnf-3β expression, whereas miR-122 indirectly inhibits c-Myc transcription by targeting Tfdp2 and E2f1. In essence, these results suggest a double-negative feedback loop between a tumor suppressor (miR-122) and an oncogene (c-Myc).
© 2013 by the American Association for the Study of Liver Diseases.
Figures
References
-
- Pasquinelli AE. MicroRNAs and their targets: recognition, regulation and an emerging reciprocal relationship. Nat Rev Genet. 2012;13:271–282. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases