

Wild type p53 transcriptionally represses the SALL2 transcription factor under genotoxic stress
- PMID: 24040083
- PMCID: PMC3765348
- DOI: 10.1371/journal.pone.0073817
Wild type p53 transcriptionally represses the SALL2 transcription factor under genotoxic stress
Erratum in
- PLoS One.2014;9(8):e104307. doi: 10.1371/journal.pone.0104307 doi: 10.1371/journal.pone.0104307
Abstract
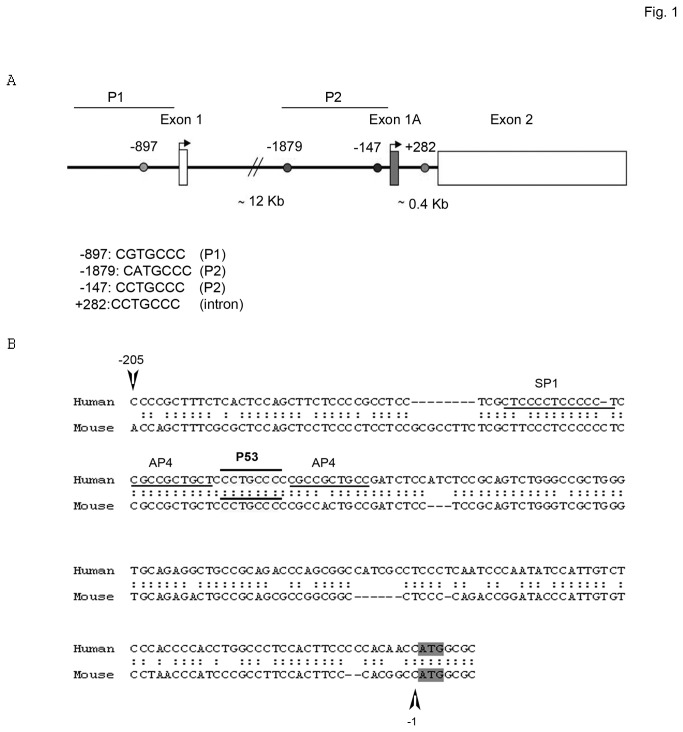
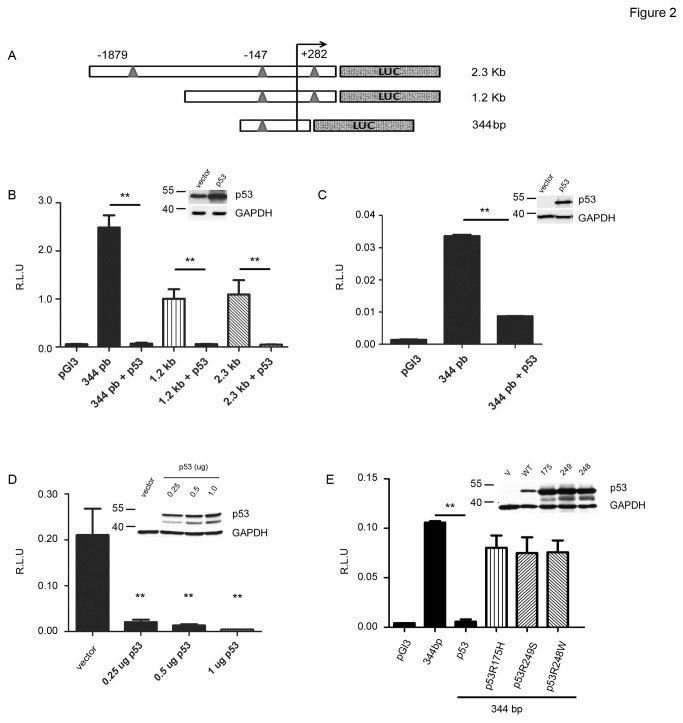
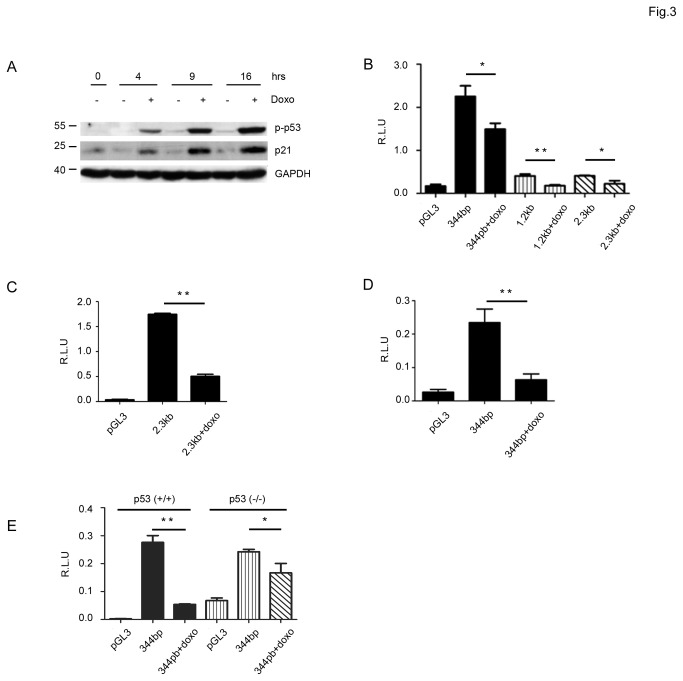
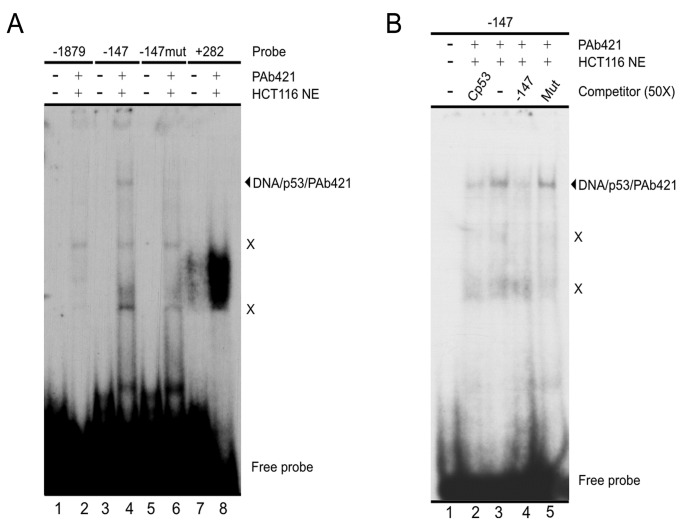
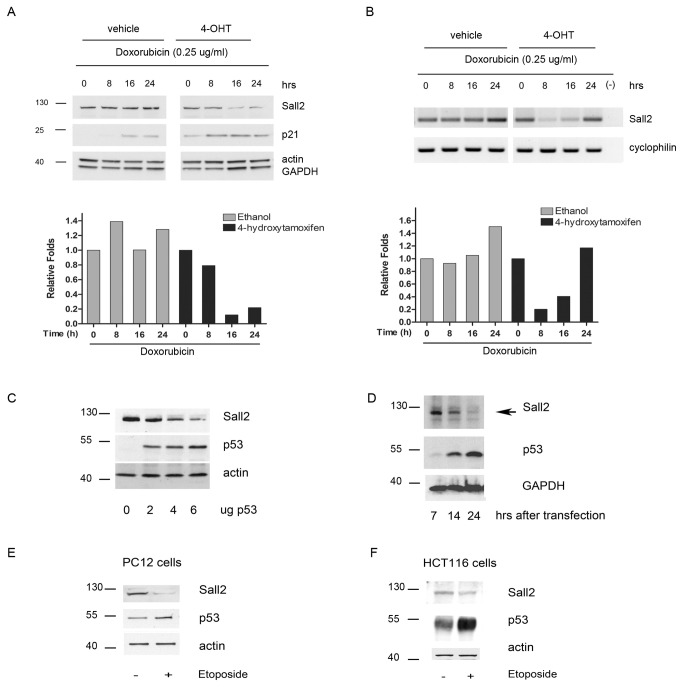
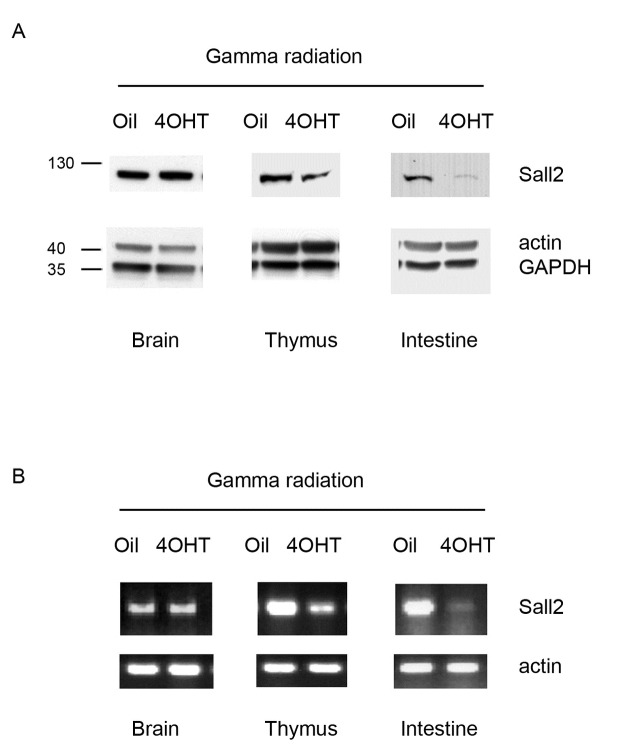
SALL2- a member of the Spalt gene family- is a poorly characterized transcription factor found deregulated in various cancers, which suggests it plays a role in the disease. We previously identified SALL2 as a novel interacting protein of neurotrophin receptors and showed that it plays a role in neuronal function, which does not necessarily explain why or how SALL2 is deregulated in cancer. Previous evidences indicate that SALL2 gene is regulated by the WT1 and AP4 transcription factors. Here, we identified SALL2 as a novel downstream target of the p53 tumor suppressor protein. Bioinformatic analysis of the SALL2 gene revealed several putative p53 half sites along the promoter region. Either overexpression of wild-type p53 or induction of the endogenous p53 by the genotoxic agent doxorubicin repressed SALL2 promoter activity in various cell lines. However R175H, R249S, and R248W p53 mutants, frequently found in the tumors of cancer patients, were unable to repress SALL2 promoter activity, suggesting that p53 specific binding to DNA is important for the regulation of SALL2. Electrophoretic mobility shift assay demonstrated binding of p53 to one of the identified p53 half sites in the Sall2 promoter, and chromatin immunoprecipitation analysis confirmed in vivo interaction of p53 with the promoter region of Sall2 containing this half site. Importantly, by using a p53ER (TAM) knockin model expressing a variant of p53 that is completely dependent on 4-hydroxy-tamoxifen for its activity, we show that p53 activation diminished SALL2 RNA and protein levels during genotoxic cellular stress in primary mouse embryo fibroblasts (MEFs) and radiosensitive tissues in vivo. Thus, our finding indicates that p53 represses SALL2 expression in a context-specific manner, adding knowledge to the understanding of SALL2 gene regulation, and to a potential mechanism for its deregulation in cancer.
Conflict of interest statement
Figures

References
-
- de Celis JF, Barrio R (2008) Regulation and function of Spalt proteins during animal development. Int J Dev Biol 52: 1-14. doi:10.1387/ijdb.072494mr. PubMed: 18033666. - DOI - PubMed
-
- Sweetman D, Münsterberg A (2006) The vertebrate spalt genes in development and disease. Dev Biol 293: 285-293. doi:10.1016/j.ydbio.2006.02.009. PubMed: 16545361. - DOI - PubMed
-
- Pincheira R, Baerwald M, Dunbar JD, Donner DB (2009) Sall2 is a novel p75NTR-interacting protein that links NGF signalling to cell cycle progression and neurite outgrowth. EMBO J 28: 261-273. doi:10.1038/emboj.2008.274. PubMed: 19131967. - DOI - PMC - PubMed
-
- Böhm J, Buck A, Borozdin W, Mannan AU, Matysiak-Scholze U et al. (2008) Sall1, sall2, and sall4 are required for neural tube closure in mice. Am J Pathol 173: 1455-1463. doi:10.2353/ajpath.2008.071039. PubMed: 18818376. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
