

Amyloid-β and APP deficiencies cause severe cerebrovascular defects: important work for an old villain
- PMID: 24040383
- PMCID: PMC3764155
- DOI: 10.1371/journal.pone.0075052
Amyloid-β and APP deficiencies cause severe cerebrovascular defects: important work for an old villain
Abstract
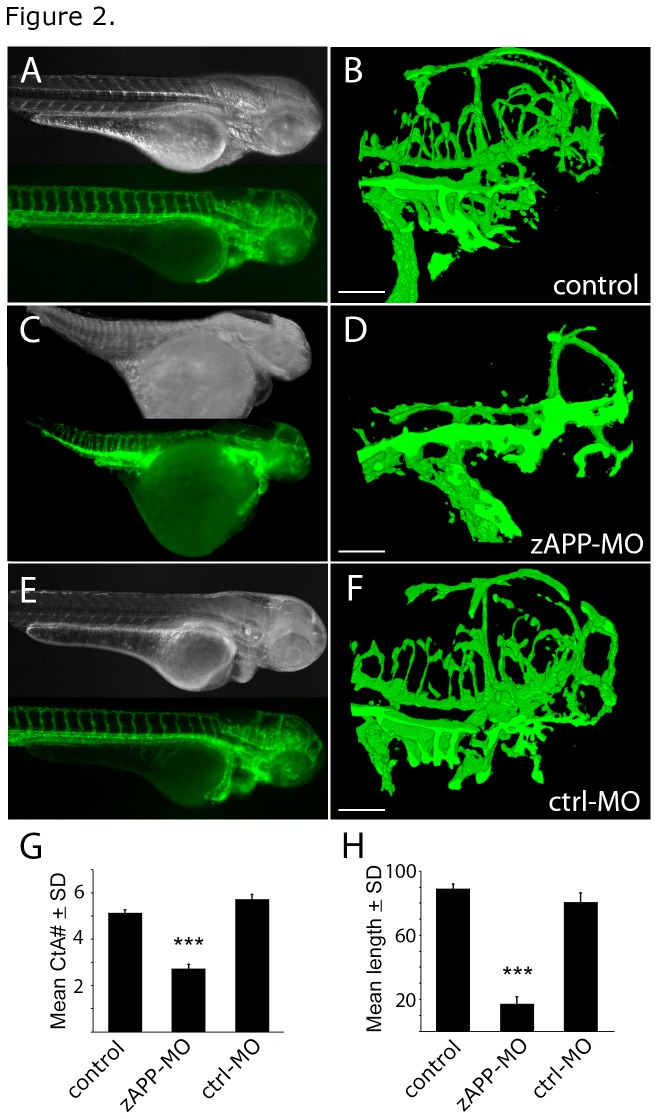
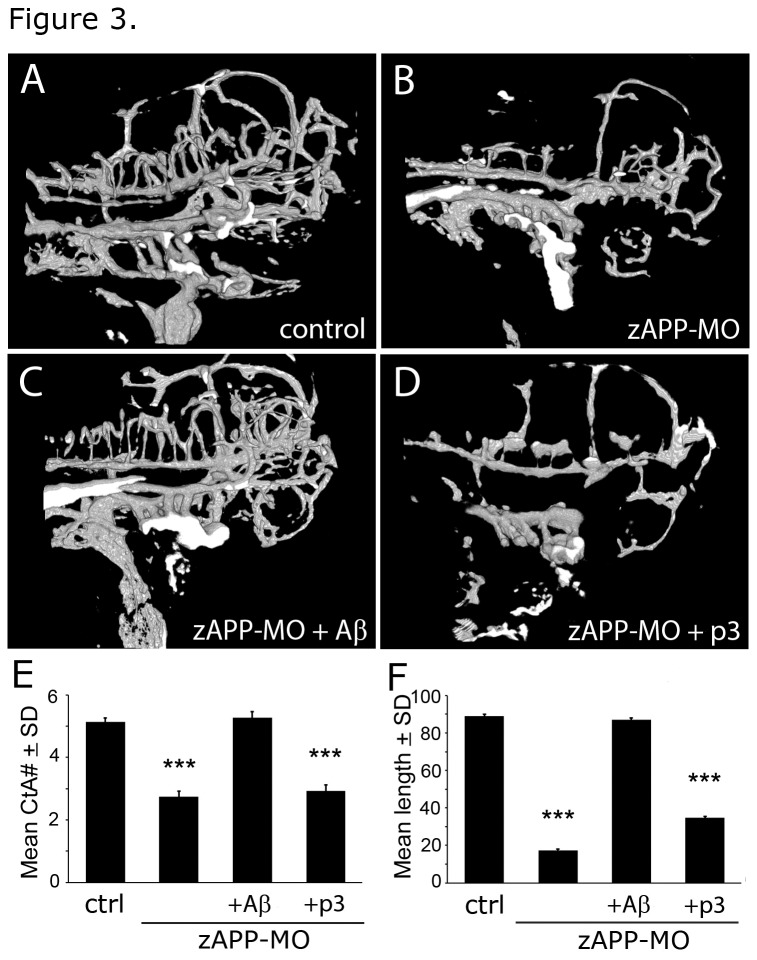
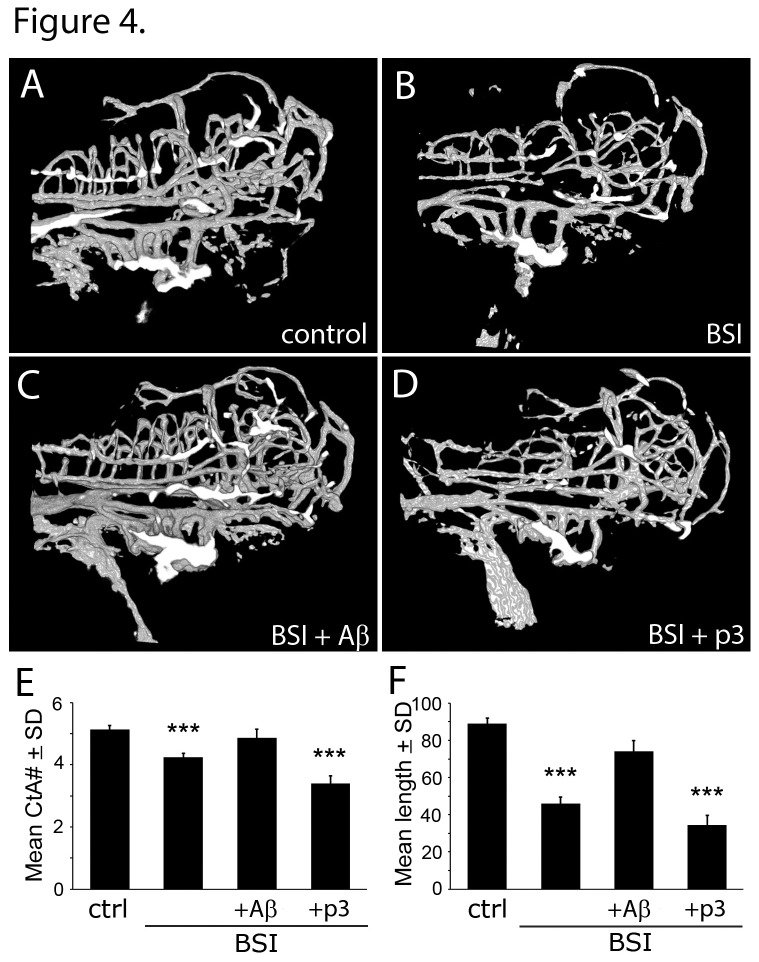
Alzheimer's disease (AD) is marked by neuritic plaques that contain insoluble deposits of amyloid-β (Aβ), yet the physiological function of this peptide has remained unclear for more than two decades. Using genetics and pharmacology we have established that Aβ plays an important role in regulating capillary bed density within the brain, a function that is distinct from other cleavage products of amyloid precursor protein (APP). APP-deficient zebrafish had fewer cerebrovascular branches and shorter vessels in the hindbrain than wild-type embryos; this phenotype was rescued by treatment with human Aβ peptide, but not a smaller APP fragment called p3. Similar vascular defects were seen in zebrafish treated with a β-secretase inhibitor (BSI) that blocked endogenous Aβ production. BSI-induced vascular defects were also improved by treatment with human Aβ, but not p3. Our results demonstrate a direct correlation between extracellular levels of Aβ and cerebrovascular density in the developing hindbrain. These findings may be relevant to AD etiology where high levels of Aβ in the brain parenchyma precede the development of neuritic plaques and dense aberrantly-branched blood vessel networks that appear between them. The ability of Aβ to modify blood vessels may coordinate capillary density with local metabolic activity, which could explain the evolutionary conservation of this peptide from lobe-finned fish to man.
Conflict of interest statement
Figures

References
-
- Theis W, Blieler L (2013) Alzheimer’s Association report: 2013 Alzheimer’s disease facts and figures. Alz Dement 9: 208-245. doi:10.1016/j.jalz.2013.02.003. - DOI - PubMed
-
- Alzheimer A (1907) Uber eine eigenartige Erkrankung der Hirnrinde. Allg Z Psychiat Psych-Gericht Med 64:146–148.
-
- Goate A, Hardy J (2012) Twenty years of Alzheimer’s disease-causing mutations. J Neurochem 120(Suppl 1): 3-8. doi:10.1111/j.1471-4159.2011.07575.x. PubMed: 22122678. - DOI - PubMed
-
- O’Brien RJ, Wong PC (2011) Amyloid precursor protein processing and Alzheimer’s disease. Annu Rev Neurosci 34: 185-204. doi:10.1146/annurev-neuro-061010-113613. PubMed: 21456963. - DOI - PMC - PubMed
-
- Cameron DJ, Galvin C, Alkam T, Sidhu H, Ellison J et al. (2012) Alzheimer’s-related peptide amyloid-β plays a conserved role in angiogenesis. PLOS ONE 7: e39598. doi:10.1371/journal.pone.0039598. PubMed: 22792182. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
